Download as pdf or txt
You might also like
- Around The World in Eighty Days: Jules VerneDocument58 pagesAround The World in Eighty Days: Jules VerneWilliam100% (3)
- Gaussian 09 User's ReferenceDocument431 pagesGaussian 09 User's ReferenceWilliamNo ratings yet
- Gaussian 09 User's ReferenceDocument431 pagesGaussian 09 User's ReferenceWilliamNo ratings yet
- Ariadne S Story Dominoes L2Document69 pagesAriadne S Story Dominoes L2William0% (1)
- Home Chiropractic HandbookDocument202 pagesHome Chiropractic HandbookFernando Bernardes78% (9)
- Summer Working Plan: AppendicesDocument11 pagesSummer Working Plan: AppendicesArshath FleminNo ratings yet
- 1260 FullDocument6 pages1260 FullAbay SisayNo ratings yet
- The Pathophysiology, Diagnosis and Treatment of Corona Virus Disease 2019 (COVID-19)Document12 pagesThe Pathophysiology, Diagnosis and Treatment of Corona Virus Disease 2019 (COVID-19)GabiiGabriielaNo ratings yet
- Cryo-Em Structure of The 2019-Ncov Spike in The Prefusion ConformationDocument2 pagesCryo-Em Structure of The 2019-Ncov Spike in The Prefusion ConformationGabriel KeresztélyNo ratings yet
- Natural Variants in Sars Cov 2 Spike Protein Pinpoint Structural and Functional Hotspots With Implications For Prophylaxis and Therapeutic StrategiesDocument10 pagesNatural Variants in Sars Cov 2 Spike Protein Pinpoint Structural and Functional Hotspots With Implications For Prophylaxis and Therapeutic StrategiesDARLENYS YADIRA MOTA MEJIANo ratings yet
- 1 s2.0 S0753332220305308 MainDocument4 pages1 s2.0 S0753332220305308 Mainvictor rodriguezNo ratings yet
- Evaluation of FlavonoidsDocument13 pagesEvaluation of Flavonoidsliem217No ratings yet
- Science Abc2241 FullDocument8 pagesScience Abc2241 FullEconomy 365No ratings yet
- 5400-Article Text-31589-2-10-20210820Document12 pages5400-Article Text-31589-2-10-20210820Voltaire Méndez RodríguezNo ratings yet
- JIANG 2021 (EPITOPO B) Epitope Profiling Reveals The Critical Antigenic Determinants in SARS-CoV-2 RBD-Based AntigenDocument14 pagesJIANG 2021 (EPITOPO B) Epitope Profiling Reveals The Critical Antigenic Determinants in SARS-CoV-2 RBD-Based AntigenJoão Pedro NunesNo ratings yet
- Neutralizing Antibodies Against SARS-CoV-2 CurrentDocument15 pagesNeutralizing Antibodies Against SARS-CoV-2 CurrentAntonius SimangunsongNo ratings yet
- Mahdian2020 Article DrugRepurposingUsingComputatioDocument9 pagesMahdian2020 Article DrugRepurposingUsingComputatioAndrew L GreearNo ratings yet
- Measuringtheserologic Responsetosevereacute Respiratorysyndrome Coronavirus2Document12 pagesMeasuringtheserologic Responsetosevereacute Respiratorysyndrome Coronavirus2Chris KontomarisNo ratings yet
- In Silico Approach To Design A Multi Epitopic Vaccine Candidate Targeting The Non Mutational Immunogenic Regions in Envelope Protein and SurfaceDocument17 pagesIn Silico Approach To Design A Multi Epitopic Vaccine Candidate Targeting The Non Mutational Immunogenic Regions in Envelope Protein and SurfacesusithraNo ratings yet
- Antiviral Research: Invited ReviewDocument16 pagesAntiviral Research: Invited ReviewasdjjjjssNo ratings yet
- Antiviral Peptides As Promising Therapeutics Against SARS-CoV 2Document8 pagesAntiviral Peptides As Promising Therapeutics Against SARS-CoV 2Stalwart sheikhNo ratings yet
- A Noncompeting Pair of Human Neutralizing Antibodies Block COVID-19 Virus Binding To Its Receptor ACE2Document6 pagesA Noncompeting Pair of Human Neutralizing Antibodies Block COVID-19 Virus Binding To Its Receptor ACE2Walid BoukhelkhalNo ratings yet
- Crystal Structure of SARS CoV 2 Nucleocapsid Protein RNA - 2020 - Acta PharmaceDocument11 pagesCrystal Structure of SARS CoV 2 Nucleocapsid Protein RNA - 2020 - Acta PharmaceMohammed Shuaib AhmedNo ratings yet
- Vaccine - RBD TArgetDocument9 pagesVaccine - RBD TArgetindahikasNo ratings yet
- Sars-Cov-2 Genetics: Key Findings For Public HealthDocument2 pagesSars-Cov-2 Genetics: Key Findings For Public HealthRishabh SachanNo ratings yet
- SARS CoV 2 Case Study A Deeper Look IntoDocument35 pagesSARS CoV 2 Case Study A Deeper Look IntogygyNo ratings yet
- Sars-Cov-2: Phylogenetic Status, Mutations and Therapeutic Research Based On Spike ProteinDocument10 pagesSars-Cov-2: Phylogenetic Status, Mutations and Therapeutic Research Based On Spike ProteinArley GutarraNo ratings yet
- ShahzaibDocument15 pagesShahzaibmanal.ghatesse5No ratings yet
- Asesoria Articulo CientificoDocument8 pagesAsesoria Articulo CientificoWilson Ramos OlanoNo ratings yet
- Coronavirus - 2020.01.30.927871.fullDocument14 pagesCoronavirus - 2020.01.30.927871.fullStephen StewartNo ratings yet
- Convalescent Blood Plasma (CBP) Donated by RecoverDocument2 pagesConvalescent Blood Plasma (CBP) Donated by RecoverMacarenaNo ratings yet
- Chloroquine study and Interaction of hydroxychloroquine with the RNA binding domain of the nucleocapsid phosphoprotein - an in silico insight into the comparative efficacy of repurposing antiviral drugsDocument14 pagesChloroquine study and Interaction of hydroxychloroquine with the RNA binding domain of the nucleocapsid phosphoprotein - an in silico insight into the comparative efficacy of repurposing antiviral drugsiran999No ratings yet
- PIIS0092867422000010Document22 pagesPIIS0092867422000010Rafaelly NascimentoNo ratings yet
- Gene of The Month: The 2019-Ncov/Sars - Cov-2 Novel Coronavirus Spike ProteinDocument4 pagesGene of The Month: The 2019-Ncov/Sars - Cov-2 Novel Coronavirus Spike ProteinNandagopal PaneerselvamNo ratings yet
- Molecular Mechanism of Evolution and Human Infection With The Novel Coronavirus (2019-Ncov)Document19 pagesMolecular Mechanism of Evolution and Human Infection With The Novel Coronavirus (2019-Ncov)Eneko AlfandegaNo ratings yet
- J Antiviral 2020 104742Document20 pagesJ Antiviral 2020 104742hanzNo ratings yet
- 409 FullDocument5 pages409 FullFacundo FrescoNo ratings yet
- The N-Glycosylation Sites and Glycan-Binding Ability of S-Protein in Sars-Cov-2 CoronavirusDocument20 pagesThe N-Glycosylation Sites and Glycan-Binding Ability of S-Protein in Sars-Cov-2 Coronaviruspopov357No ratings yet
- Covid Reseptor ACEDocument19 pagesCovid Reseptor ACETitik HidayatiNo ratings yet
- Caractere VirologiaueDocument29 pagesCaractere Virologiauehind el hamriNo ratings yet
- Journal of Infection and Public HealthDocument10 pagesJournal of Infection and Public Healthtuanda222No ratings yet
- Prediction and Analysis of Key Protein Structures of 2019-NcovDocument10 pagesPrediction and Analysis of Key Protein Structures of 2019-NcovGiuseppe GrecoNo ratings yet
- Mapping The Human Genetic Architecture of COVID-19: ArticleDocument39 pagesMapping The Human Genetic Architecture of COVID-19: ArticleZara FustaNo ratings yet
- WissenshaftDocument11 pagesWissenshaftkacagj bajazzoNo ratings yet
- Virology: SciencedirectDocument8 pagesVirology: Sciencedirectathoillah_JrNo ratings yet
- nCoV - Med - 2020-06-05 - Enhanced Receptor Binding of SARS-CoV-2 Through Networks of Hydrogen-Bonding and Hydrophobic InteractionsDocument8 pagesnCoV - Med - 2020-06-05 - Enhanced Receptor Binding of SARS-CoV-2 Through Networks of Hydrogen-Bonding and Hydrophobic InteractionsKaroline MarxNo ratings yet
- Natrulazing AntibodiDocument6 pagesNatrulazing Antibodidita novia maharaniNo ratings yet
- Virus Research: A B A B B A B A B ADocument9 pagesVirus Research: A B A B B A B A B AKEEP CALM studiosNo ratings yet
- Journal Pre-Proof: Infection, Genetics and EvolutionDocument29 pagesJournal Pre-Proof: Infection, Genetics and EvolutionMohammed Shuaib AhmedNo ratings yet
- Structure-Based Drug Discovery To Identify SARS-CoV2 Spike Protein ACE2 Interaction InhibitorsDocument20 pagesStructure-Based Drug Discovery To Identify SARS-CoV2 Spike Protein ACE2 Interaction Inhibitorsdenemebakalim72No ratings yet
- 1 s2.0 S096999612030406X MainDocument12 pages1 s2.0 S096999612030406X MainLuis ricardo Véjar felixNo ratings yet
- Jurnal Sinovac 2Document6 pagesJurnal Sinovac 2Delapan SembilanNo ratings yet
- 2022 - ACE2 SusceptibilidadDocument13 pages2022 - ACE2 SusceptibilidadMario Martínez TorijaNo ratings yet
- Overwhelming Mutations or SNPs of SARS-CoV-2: A Point of CautionDocument4 pagesOverwhelming Mutations or SNPs of SARS-CoV-2: A Point of CautionDanilo RosaNo ratings yet
- Sars-Cov-2 Receptor and Renin-Angiotensin System Regulation: Impact of Genetics Variants in Ace2 GeneDocument9 pagesSars-Cov-2 Receptor and Renin-Angiotensin System Regulation: Impact of Genetics Variants in Ace2 GeneInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- Chen Et AlDocument6 pagesChen Et AlHarika PutraNo ratings yet
- Grupo01 QYF 2023Document6 pagesGrupo01 QYF 2023Fernanda ZapataNo ratings yet
- Current Research in Pharmacology and Drug Discovery: Pro Pro ProDocument23 pagesCurrent Research in Pharmacology and Drug Discovery: Pro Pro ProZac IngNo ratings yet
- Imun ResponseDocument9 pagesImun ResponsePena NofaNo ratings yet
- Covid Update 2021Document30 pagesCovid Update 2021Andreea PostolacheNo ratings yet
- COVID-19 Gain of FunctionDocument5 pagesCOVID-19 Gain of FunctionRed Cloud ShamanNo ratings yet
- Corona VirusDocument22 pagesCorona Virusmaria carolina olazabal rodriguezNo ratings yet
- Drug Sars-CovDocument20 pagesDrug Sars-CovSilvi Zakiyatul IlmiyahNo ratings yet
- Structural and Functional Basis of Sars-Cov-2 Entry by Using Human Ace2Document21 pagesStructural and Functional Basis of Sars-Cov-2 Entry by Using Human Ace2rezaNo ratings yet
- Viruses: Sars-Cov-2 Spike Impairs Dna Damage Repair and Inhibits V (D) J Recombination in VitroDocument10 pagesViruses: Sars-Cov-2 Spike Impairs Dna Damage Repair and Inhibits V (D) J Recombination in Vitromuun yayo100% (1)
- DYMO SDK For Linux - CUPS DriversDocument5 pagesDYMO SDK For Linux - CUPS DriversWilliamNo ratings yet
- The Vesuvius Mosaic Dominoes Level 3Document44 pagesThe Vesuvius Mosaic Dominoes Level 3WilliamNo ratings yet
- Tristan and Isolde PDFDocument52 pagesTristan and Isolde PDFWilliamNo ratings yet
- Multiconfigurational and Multireference Methods: Peter R. Taylor and Jeppe OlsenDocument55 pagesMulticonfigurational and Multireference Methods: Peter R. Taylor and Jeppe OlsenWilliamNo ratings yet
- Introduction To Second-Quantization IDocument16 pagesIntroduction To Second-Quantization IWilliamNo ratings yet
- Introduction To Second-Quantization IIDocument16 pagesIntroduction To Second-Quantization IIWilliamNo ratings yet
- Test Grammar 3Document6 pagesTest Grammar 3Veronika EneliaNo ratings yet
- 1 - ATP-ADP - CycleDocument10 pages1 - ATP-ADP - CycleYay SandovalNo ratings yet
- Acupuncture IVF by DR Lifang Liang PDFDocument226 pagesAcupuncture IVF by DR Lifang Liang PDFNick Max100% (1)
- Detailed Lesson Plan in Science 6: Second QuarterDocument5 pagesDetailed Lesson Plan in Science 6: Second QuarterAngel R. CortesNo ratings yet
- Respiration in PlantsDocument13 pagesRespiration in Plantsc7zb7hhq64No ratings yet
- BiologikaOrganAtlas2021 en BasicsDocument34 pagesBiologikaOrganAtlas2021 en Basicskadarzoltan75% (4)
- Theories of Crime Early Biological TheoriesDocument6 pagesTheories of Crime Early Biological TheoriesCharmis TubilNo ratings yet
- AnabolismeDocument27 pagesAnabolismeRofina PutriNo ratings yet
- FCI Previous Question PapersDocument5 pagesFCI Previous Question PapersRohit BeriNo ratings yet
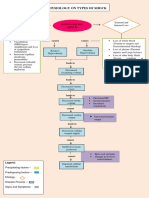
- Pathophysiology On Types of ShockDocument4 pagesPathophysiology On Types of ShockJessa Mae Alforque AsentistaNo ratings yet
- Abegg, Entiauspe-Neto, de Lema - 2015 - First Record of Albinism in The Elapomorphini Tribe (Serpentes Dipsadidae)Document4 pagesAbegg, Entiauspe-Neto, de Lema - 2015 - First Record of Albinism in The Elapomorphini Tribe (Serpentes Dipsadidae)Diego Gomiero CavalheriNo ratings yet
- CalciumsDocument5 pagesCalciums786lailaNo ratings yet
- Biophysical Chemistry of The Ageing Eye Lens.: Proquest Document LinkDocument5 pagesBiophysical Chemistry of The Ageing Eye Lens.: Proquest Document Linkmau tauNo ratings yet
- The Forsaken FrontierDocument137 pagesThe Forsaken FrontierpverdanceNo ratings yet
- Yds Deneme Sinavi PDFDocument16 pagesYds Deneme Sinavi PDFodysseyyyNo ratings yet
- 2nd PUC QUESTION PAPERS Chemistry 2006-2010Document21 pages2nd PUC QUESTION PAPERS Chemistry 2006-2010Mohan Kumar PNo ratings yet
- Citric AcidDocument7 pagesCitric AcidNisal PanagodaNo ratings yet
- Summary of Pub Med Articles Protandim PDFDocument2 pagesSummary of Pub Med Articles Protandim PDFLeslee MasonNo ratings yet
- Major Examination: Subject - Science SESSION 2019-20 Maximum Marks: 80 Class - VII Time: 3 HoursDocument22 pagesMajor Examination: Subject - Science SESSION 2019-20 Maximum Marks: 80 Class - VII Time: 3 HoursCosmos WithmeNo ratings yet
- Bio Paper FinalDocument8 pagesBio Paper FinalNiem PhamNo ratings yet
- Carla Hernandez 100Document4 pagesCarla Hernandez 100JhunnieEy ReyesNo ratings yet
- Kebo101 MergedDocument252 pagesKebo101 Mergedik3452etNo ratings yet
- Karyotype of Native Zea Mays Ssp. Mays Proto-Confite MorochoDocument6 pagesKaryotype of Native Zea Mays Ssp. Mays Proto-Confite MorochoLilian CarcamoNo ratings yet
- Anatomy of Urinary Tract Systems - 20240117 - 125604 - 0000Document44 pagesAnatomy of Urinary Tract Systems - 20240117 - 125604 - 0000Sneha ThiagarajanNo ratings yet
- Generation and Harvesting of Microalgae Biomass For Biofuel ProductionDocument23 pagesGeneration and Harvesting of Microalgae Biomass For Biofuel ProductionFábio UcellaNo ratings yet
- Attention and Concentration in SportsDocument30 pagesAttention and Concentration in SportsArundhathy GopakumarNo ratings yet
- Review Metabolit Sekunder Dan Aktivitas FarmakologDocument8 pagesReview Metabolit Sekunder Dan Aktivitas FarmakologAnastasia ElisaNo ratings yet
- Ankylosing SpondylitisDocument19 pagesAnkylosing SpondylitisZulhida YuniNo ratings yet