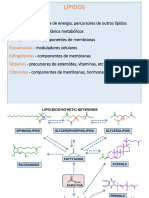
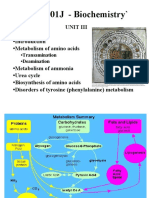
Download as pdf or txt
You might also like
- List of Books PDFDocument29 pagesList of Books PDFDevansh Agarwal75% (4)
- Regulation 391ADocument6 pagesRegulation 391Adnageshm4n244100% (1)
- Acetyl Coenzyme A - AssignmentDocument2 pagesAcetyl Coenzyme A - AssignmentSasa LiliNo ratings yet
- Ketosis-Causes AND Consequences: Biochemistry For Medics WWW - Namrata.coDocument39 pagesKetosis-Causes AND Consequences: Biochemistry For Medics WWW - Namrata.corohishaakNo ratings yet
- Troduction:: Ketone Bodies Are Three Chemicals That Are Produced When Fatty Acids Are BrokenDocument11 pagesTroduction:: Ketone Bodies Are Three Chemicals That Are Produced When Fatty Acids Are BrokenMshoaibNo ratings yet
- Lecture 4 Ketone Body MetabolismDocument239 pagesLecture 4 Ketone Body MetabolismNANDINI RATTANNo ratings yet
- Amino Acid Oxidation and The Production PDFDocument34 pagesAmino Acid Oxidation and The Production PDFNini BesiNo ratings yet
- Amino Acid Metabolism 2Document29 pagesAmino Acid Metabolism 2Francis GacheruNo ratings yet
- Acetyl CoADocument2 pagesAcetyl CoAkjsldfkjasdlfkNo ratings yet
- Fat SynthesisDocument42 pagesFat Synthesisjn724544No ratings yet
- Amino Acids OverviewDocument30 pagesAmino Acids OverviewbrownhazelNo ratings yet
- Biosintesis Asam Lemak 1Document32 pagesBiosintesis Asam Lemak 1YainPanggaloNo ratings yet
- Ehninger Principles of Biochemistry, Fourth Edition - DavidNelson, Michael M. Cox (0641-0680) PDFDocument40 pagesEhninger Principles of Biochemistry, Fourth Edition - DavidNelson, Michael M. Cox (0641-0680) PDFDesmon Jonathan SumolangNo ratings yet
- Fatty Acid SynthesisDocument3 pagesFatty Acid SynthesisClairyssa Myn D CaballeroNo ratings yet
- Notes On AminoAcid MetabolismDocument13 pagesNotes On AminoAcid MetabolismMaya Abou MradNo ratings yet
- Lipid MetabolismDocument6 pagesLipid Metabolismfollower 2No ratings yet
- Fatty Acid BiosynthesisDocument7 pagesFatty Acid BiosynthesisJeremiah Eyo AmanamNo ratings yet
- Metabolism of Lipids (2) NewDocument64 pagesMetabolism of Lipids (2) NewLyra Get100% (1)
- C10 Protein and Amino Acid MetabolismDocument8 pagesC10 Protein and Amino Acid MetabolismSoraya D. Al-ObinayNo ratings yet
- BCH 1212 Lipid Metabolism 1Document44 pagesBCH 1212 Lipid Metabolism 1Hamirie JoshuaNo ratings yet
- Lipid Metabolism: Metabolism of Fatty Acids - IDocument24 pagesLipid Metabolism: Metabolism of Fatty Acids - Icheckmate100% (1)
- Amphibolic Nature of Krebs Cycle: How What We Are Is What We EatDocument33 pagesAmphibolic Nature of Krebs Cycle: How What We Are Is What We EatSoloNo ratings yet
- Zhang Et Al. - 2021 - Metabolism and Strategies For Enhanced Supply of ADocument14 pagesZhang Et Al. - 2021 - Metabolism and Strategies For Enhanced Supply of AAnaNo ratings yet
- Citric Acid CycleDocument2 pagesCitric Acid CyclePablo MaldonadoNo ratings yet
- BIOSINTESISDocument34 pagesBIOSINTESISAyodia RanggiNo ratings yet
- Lipid Metabolism LASTDocument80 pagesLipid Metabolism LASTAhmed JamalNo ratings yet
- Bch-Ii 28032022Document11 pagesBch-Ii 28032022Hawaid AhmadNo ratings yet
- CHAPTER 24 Lipid BiosynthesisDocument16 pagesCHAPTER 24 Lipid Biosynthesis楊畯凱No ratings yet
- Biosynthesis of Fatty AcidsDocument3 pagesBiosynthesis of Fatty AcidsAhsan AliNo ratings yet
- Ketone Metabolism by Savath SovannakDocument5 pagesKetone Metabolism by Savath SovannakRith Hengmeng (Study Smart)No ratings yet
- Biochem Rev Chap 8Document3 pagesBiochem Rev Chap 8Jacob PallorinaNo ratings yet
- FA SynthesisDocument12 pagesFA SynthesisRith Hengmeng (Study Smart)No ratings yet
- The Krebs Cycle: Removal of Hydrogens and Electrons, and The Release of CODocument24 pagesThe Krebs Cycle: Removal of Hydrogens and Electrons, and The Release of COLeann RodriguezNo ratings yet
- Introduction Amino Acid Matabolism and CatabolismDocument45 pagesIntroduction Amino Acid Matabolism and CatabolismAboubakar Moalim Mahad moh'd100% (1)
- 2 Aula 21.22 Síntese de Ácidos GordosDocument42 pages2 Aula 21.22 Síntese de Ácidos GordosFilipa MendesNo ratings yet
- Biosynthesis Fatty AcidsDocument59 pagesBiosynthesis Fatty AcidsKate Alyssa Caton100% (1)
- Lipid Metabolism 2023Document36 pagesLipid Metabolism 2023Moses MutsikwiNo ratings yet
- Chapter 23 Biosynthesis of Fatty Acids - EicosanoidsDocument3 pagesChapter 23 Biosynthesis of Fatty Acids - Eicosanoidst4gjzhpfjcNo ratings yet
- M - 29 Lipid MetabolismDocument5 pagesM - 29 Lipid MetabolismDr. Tapan Kr. DuttaNo ratings yet
- 10.1016@0968 00048690256 2Document3 pages10.1016@0968 00048690256 2nnbfgwnrhbNo ratings yet
- Protein MetabolismDocument19 pagesProtein MetabolismAnn Michelle Tarrobago100% (1)
- Lipid MetabolismDocument28 pagesLipid Metabolismperssivesimangavwa2002No ratings yet
- 11004-Article Text PDF-31187-2-10-20180516Document12 pages11004-Article Text PDF-31187-2-10-20180516BrianWirawanNo ratings yet
- Ketogenesis and KetolysisDocument4 pagesKetogenesis and KetolysisSergio D. Sánchez VillamarínNo ratings yet
- Aa & Nucleotide MetabolismDocument34 pagesAa & Nucleotide MetabolismEvaNo ratings yet
- Lipid Metabolism: Stoker Chapter 25 Lippincott Chapter 16Document21 pagesLipid Metabolism: Stoker Chapter 25 Lippincott Chapter 16Shane G.No ratings yet
- Citric Acid Cycle PDFDocument26 pagesCitric Acid Cycle PDFjairajNo ratings yet
- Sintesis LemakDocument35 pagesSintesis LemakAndriati RahayuNo ratings yet
- Ketogenesis and KetolysisDocument4 pagesKetogenesis and KetolysisSergio D. Sánchez VillamarínNo ratings yet
- Fatty Acid SynthesisDocument38 pagesFatty Acid SynthesisEmm NomanNo ratings yet
- Item-9,10, Lipid MetabolismDocument38 pagesItem-9,10, Lipid MetabolismSheikh FahadNo ratings yet
- 19CACCM ProvidoDocument1 page19CACCM ProvidoYUAN PROVIDONo ratings yet
- Acetic Acid: 2 PropertiesDocument12 pagesAcetic Acid: 2 PropertiesGudang MyosiNo ratings yet
- Unit 9 Lipid MetabolismDocument28 pagesUnit 9 Lipid MetabolismAthina100% (1)
- Biosynthesis of Saturated Fatty Acid 111Document10 pagesBiosynthesis of Saturated Fatty Acid 111ThvNo ratings yet
- Biochemistry Unit 03 PDFDocument66 pagesBiochemistry Unit 03 PDFDhanush kannanNo ratings yet
- Metabolism OF Amino Acids: Yulia SuciatiDocument44 pagesMetabolism OF Amino Acids: Yulia SuciatinrsNo ratings yet
- AST and ALTDocument7 pagesAST and ALTTrisha NavarceNo ratings yet
- Fatty Acid SynthesisDocument28 pagesFatty Acid Synthesishassanainshahi13No ratings yet
- Amino Acid Metabolism All LecturesDocument18 pagesAmino Acid Metabolism All Lecturesmizare29gNo ratings yet
- Ketogenesis and KetolysisDocument17 pagesKetogenesis and KetolysisValine Cysteine Methionine100% (2)
- Neural Responses Associated With Positive and Negative Emotion Processing in Patients With Left Versus Right Temporal Lobe EpilepsyDocument9 pagesNeural Responses Associated With Positive and Negative Emotion Processing in Patients With Left Versus Right Temporal Lobe EpilepsyRuth Noemy Ruiz MangandiNo ratings yet
- Marlow 1991Document3 pagesMarlow 1991Ruth Noemy Ruiz MangandiNo ratings yet
- Compacted Silage Fermentation On Whole Sweet Sorghum Plant For Distributed Bioethanol ProductionDocument8 pagesCompacted Silage Fermentation On Whole Sweet Sorghum Plant For Distributed Bioethanol ProductionRuth Noemy Ruiz MangandiNo ratings yet
- Bred Dam 1991Document7 pagesBred Dam 1991Ruth Noemy Ruiz MangandiNo ratings yet
- Baba Yan 1981Document6 pagesBaba Yan 1981Ruth Noemy Ruiz MangandiNo ratings yet
- Comparison of Key Odorants Generated by Thermal Treatment of Commercial and Self-Prepared Yeast Extracts: Influence of The Amino Acid Composition On Odorant FormationDocument7 pagesComparison of Key Odorants Generated by Thermal Treatment of Commercial and Self-Prepared Yeast Extracts: Influence of The Amino Acid Composition On Odorant FormationRuth Noemy Ruiz MangandiNo ratings yet
- Approach To Microalgal Biodiesel Production: Insight Review On Recent Advancements and Future OutlookDocument19 pagesApproach To Microalgal Biodiesel Production: Insight Review On Recent Advancements and Future OutlookRuth Noemy Ruiz MangandiNo ratings yet
- Pareyt 2010Document8 pagesPareyt 2010Ruth Noemy Ruiz MangandiNo ratings yet
- Hemmati2021 PDFDocument16 pagesHemmati2021 PDFRuth Noemy Ruiz MangandiNo ratings yet
- Use of Coffee Waste For The Production of BiofuelsDocument9 pagesUse of Coffee Waste For The Production of BiofuelsRuth Noemy Ruiz MangandiNo ratings yet
- Prevention of Discoloration in Retorted Whole Egg: A Research NoteDocument2 pagesPrevention of Discoloration in Retorted Whole Egg: A Research NoteRuth Noemy Ruiz MangandiNo ratings yet
- Eastlake2002 PDFDocument9 pagesEastlake2002 PDFRuth Noemy Ruiz MangandiNo ratings yet
- Prevention of The Green-Gray Discoloration in Cooked Liquid Whole EggsDocument4 pagesPrevention of The Green-Gray Discoloration in Cooked Liquid Whole EggsRuth Noemy Ruiz MangandiNo ratings yet
- AscoliteDocument3 pagesAscoliteAntony Valentine PatrickNo ratings yet
- Chemical Bonds and Molecular StructureDocument2 pagesChemical Bonds and Molecular StructureAtlas CruxNo ratings yet
- Of Electrons That An Atom Either Gains or Loses in Order To Form A Chemical Bond With Another Atom. Example: MN 2+, O 2-, Fe 2+, CL - EtcDocument5 pagesOf Electrons That An Atom Either Gains or Loses in Order To Form A Chemical Bond With Another Atom. Example: MN 2+, O 2-, Fe 2+, CL - EtcAsif FarhanNo ratings yet
- Manufacturing Industry ReportDocument3 pagesManufacturing Industry ReportShubham SayreNo ratings yet
- Muscular Development - June 2017Document164 pagesMuscular Development - June 2017Luís Gomes100% (1)
- Acroleina Agua AzeotropoDocument1 pageAcroleina Agua AzeotropoAnonymous ZAr1RKNo ratings yet
- HTV Sh5070u MSDS (E) PDFDocument5 pagesHTV Sh5070u MSDS (E) PDFAnie WajidNo ratings yet
- 0625 w18 QP 23Document16 pages0625 w18 QP 23saeed akhtarNo ratings yet
- Week 2 Roxas Read and AnswerDocument11 pagesWeek 2 Roxas Read and AnswerPrecious Nichole FloridaNo ratings yet
- Natural Toxins: 1. Toxic Phenolic SubstancesDocument12 pagesNatural Toxins: 1. Toxic Phenolic SubstancesMohamed Bayoumi mohamed100% (1)
- Reading Comprehension: SubjectsDocument27 pagesReading Comprehension: SubjectsRikeNo ratings yet
- Carbon Fiber Prepreg PDSDocument1 pageCarbon Fiber Prepreg PDSMauro RosarioNo ratings yet
- Synthesis of General Distillation Sequences-Nonsharp SeparationsDocument23 pagesSynthesis of General Distillation Sequences-Nonsharp SeparationsJorge Mario RodriguezNo ratings yet
- Hydraulic and Pneumatic ApplicationDocument34 pagesHydraulic and Pneumatic Applicationjhon jairo arangoNo ratings yet
- Material Safety Data Sheet: 1. Product and CompanyDocument5 pagesMaterial Safety Data Sheet: 1. Product and CompanyJezrell JaravataNo ratings yet
- Textbook of Forensic PharmacyDocument201 pagesTextbook of Forensic PharmacyAmbika DurgNo ratings yet
- Maintenance of Electrodes EnglishDocument2 pagesMaintenance of Electrodes EnglishLaboratorium Lingkungan Tulungagung100% (1)
- TM Prepare & Produce Cakes & Pastries FinalDocument136 pagesTM Prepare & Produce Cakes & Pastries FinalNor AzurawatiNo ratings yet
- Siliconizing Process of Mild Steel Substrate by Using Tronoh Si - 2015 - Procedi PDFDocument6 pagesSiliconizing Process of Mild Steel Substrate by Using Tronoh Si - 2015 - Procedi PDFMohd TariqNo ratings yet
- Final Year Project:: Engr. Hasan JehanzaibDocument56 pagesFinal Year Project:: Engr. Hasan JehanzaibMuhammad AbdullahNo ratings yet
- International - Standard ISO 14175Document3 pagesInternational - Standard ISO 14175Geom_MengoliNo ratings yet
- Differences in Chemical Composition and Digestibility of Beet and Cane MolassesDocument9 pagesDifferences in Chemical Composition and Digestibility of Beet and Cane MolassesGading HerinnayahNo ratings yet
- Air Pump2Document44 pagesAir Pump2abduallah muhammadNo ratings yet
- Mapua Institute of Technology Department of Physics: Grade GradesDocument4 pagesMapua Institute of Technology Department of Physics: Grade GradesMichaela VillaflorNo ratings yet
- Revealing PAGBsDocument61 pagesRevealing PAGBsmalevolentNo ratings yet
- M.pharm. Quality Assurance SyllabusDocument19 pagesM.pharm. Quality Assurance SyllabusDang Anh DuyNo ratings yet
- SPM Biology Notes - NutritionDocument4 pagesSPM Biology Notes - NutritionSPM50% (2)