
BBB 0007
BBB 0007
You might also like
- Biofilms in Treatment of Waste Water 2Document15 pagesBiofilms in Treatment of Waste Water 2anishapradhan.studNo ratings yet
- Engineering Biofilm Formation and Dispersal: Thomas K. Wood, Seok Hoon Hong and Qun MaDocument8 pagesEngineering Biofilm Formation and Dispersal: Thomas K. Wood, Seok Hoon Hong and Qun MaShampa SenNo ratings yet
- Kishen - Haapasalo - 2010 - Biofilm Models and Methods of Biofilm AssessmentDocument21 pagesKishen - Haapasalo - 2010 - Biofilm Models and Methods of Biofilm AssessmentdrchanyccNo ratings yet
- Corrosion Mic &mitigationDocument6 pagesCorrosion Mic &mitigationprakashNo ratings yet
- Biofilm Models and Methods ofDocument21 pagesBiofilm Models and Methods ofVerghese George MNo ratings yet
- 2010 79 711 FEOFILOVA Fungal Cell Wall MicrobiolDocument10 pages2010 79 711 FEOFILOVA Fungal Cell Wall MicrobiolGebrehiwot GebremedhinNo ratings yet
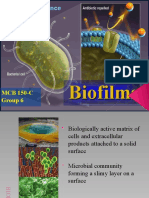
- MCB 150-C Group 6Document31 pagesMCB 150-C Group 6Angela Kristel DalaoNo ratings yet
- Https Webmail - Med.auth 1Document10 pagesHttps Webmail - Med.auth 1DD DNo ratings yet
- Microcolonies StructureDocument12 pagesMicrocolonies StructuremelanypaguayNo ratings yet
- Fleming and Wingender Biofilm Matrix Nature ReviewsDocument11 pagesFleming and Wingender Biofilm Matrix Nature Reviewsgkhan3No ratings yet
- Vol. 6, Issue 1, January 2018, PharmaTutor, Paper-2Document7 pagesVol. 6, Issue 1, January 2018, PharmaTutor, Paper-2mmmmmNo ratings yet
- Plasma Activated WaterDocument12 pagesPlasma Activated WaterconsendoNo ratings yet
- Biofilms in Endodontic InfectionsDocument10 pagesBiofilms in Endodontic InfectionsValderrama Cisneros José LuisNo ratings yet
- Biofilms and Their Advantages/Disadvantages in Food IndustryDocument7 pagesBiofilms and Their Advantages/Disadvantages in Food IndustryUmarNo ratings yet
- Biodegradacion de XenobioticosDocument8 pagesBiodegradacion de XenobioticosDoc Efra RamirezNo ratings yet
- Biological Fixed Film: Md. Shahinoor Islam, Geelsu Hwang, Yang LiuDocument33 pagesBiological Fixed Film: Md. Shahinoor Islam, Geelsu Hwang, Yang Liusong1088No ratings yet
- Bharti2017 Phototrophic BiofilmsDocument16 pagesBharti2017 Phototrophic BiofilmsSacra PsyntergiaNo ratings yet
- BIOFILMDocument79 pagesBIOFILMrkc456No ratings yet
- Reviews: Bacterial Biofilms: From The Natural Environment To Infectious DiseasesDocument15 pagesReviews: Bacterial Biofilms: From The Natural Environment To Infectious DiseasesJordy KunNo ratings yet
- Sujit - Cife - Role of Biofilm in Aquaculture - FinalDocument55 pagesSujit - Cife - Role of Biofilm in Aquaculture - FinalsushanfcriNo ratings yet
- Biofilm SemDocument64 pagesBiofilm SemmaltiNo ratings yet
- 250 Flemming Wingender ThebiofilmmatrixDocument12 pages250 Flemming Wingender ThebiofilmmatrixdeviimaNo ratings yet
- Chapter 2. Biotechnology Principles of Solid State FermentationDocument52 pagesChapter 2. Biotechnology Principles of Solid State Fermentationm.alejandracruzsalazarNo ratings yet
- InTech-Scanning Electron Microscopy Sem and Environmental Sem Suitable Tools For Study of Adhesion Stage and Biofilm FormationDocument15 pagesInTech-Scanning Electron Microscopy Sem and Environmental Sem Suitable Tools For Study of Adhesion Stage and Biofilm FormationJose Manuel Oliveira PereiraNo ratings yet
- Galley Proof Ms Ijbpas 2021 5610Document9 pagesGalley Proof Ms Ijbpas 2021 5610saisree malladiNo ratings yet
- Biofilm FormationDocument12 pagesBiofilm FormationFhaliesha ZakurNo ratings yet
- Biofilms: Microbial Life On Surfaces: Rodney M. DonlanDocument10 pagesBiofilms: Microbial Life On Surfaces: Rodney M. DonlanKapil JhajhariaNo ratings yet
- Maslowski 2019 Metabolism at The Centre Clinical and Experimental ImmunologyDocument13 pagesMaslowski 2019 Metabolism at The Centre Clinical and Experimental ImmunologyMarcelo MurilloNo ratings yet
- The Soil Microbiome - From Metagenomics To Metaphenomics: SciencedirectDocument7 pagesThe Soil Microbiome - From Metagenomics To Metaphenomics: SciencedirectMiroz RexNo ratings yet
- The Superficial Life of MicrobesDocument3 pagesThe Superficial Life of MicrobesHarveenkaur MalhanNo ratings yet
- Articulo Grupo1Document17 pagesArticulo Grupo1PAULA ANDREA QUICENO GIRALDONo ratings yet
- Bio Film: Emilia Oleandro, Romina Rega, Martina Mugnano, Filomena Nazzaro, Pietro Ferraro, Simonetta GrilliDocument10 pagesBio Film: Emilia Oleandro, Romina Rega, Martina Mugnano, Filomena Nazzaro, Pietro Ferraro, Simonetta GrilliPFENo ratings yet
- Biofilm ReportDocument50 pagesBiofilm ReportMary Joy DalisayNo ratings yet
- Detection of pathogenesis Related proteins- chitinase and β-1,3-glucanase in induced chickpeaDocument4 pagesDetection of pathogenesis Related proteins- chitinase and β-1,3-glucanase in induced chickpeaImmu JiNo ratings yet
- Vanysacker-2013-Microbial Adhesion and BiofilmDocument12 pagesVanysacker-2013-Microbial Adhesion and BiofilmEduard-Marius LungulescuNo ratings yet
- Fmicb 12 675843Document15 pagesFmicb 12 675843Rin ChanNo ratings yet
- Microbiology and Biotechnology ScriptDocument18 pagesMicrobiology and Biotechnology ScriptDavid RăscuţoiNo ratings yet
- CHEMOSTATDocument12 pagesCHEMOSTATHass AnNo ratings yet
- Surface ChemistryDocument15 pagesSurface ChemistryHajime HikariNo ratings yet
- Why Do Microbes Make MineralsDocument40 pagesWhy Do Microbes Make MineralsVelraj ParthibanNo ratings yet
- Tissue Engineering 4Document20 pagesTissue Engineering 4Vidit NarayanNo ratings yet
- Nghien CuuDocument9 pagesNghien CuuNguyen GiaoNo ratings yet
- Biofilm Dispersion: Kendra P. Rumbaugh and Karin SauerDocument16 pagesBiofilm Dispersion: Kendra P. Rumbaugh and Karin SauerTallesNo ratings yet
- Hamedi 2017Document22 pagesHamedi 2017joaovictorandre0609No ratings yet
- Content 9080Document7 pagesContent 9080Andreea MicuNo ratings yet
- Review AdhesionDocument16 pagesReview AdhesionMirella DannaNo ratings yet
- Bio FilmDocument7 pagesBio FilmFREIMUZICNo ratings yet
- Bertrand Et Al. 2021 - Estimation of Pore Dimensions in Lipid Membranes Induced by Peptides and Other Biomolecules A ReviewDocument18 pagesBertrand Et Al. 2021 - Estimation of Pore Dimensions in Lipid Membranes Induced by Peptides and Other Biomolecules A ReviewAmadorRevillaNo ratings yet
- Content ServerDocument10 pagesContent ServerAdit VekariaNo ratings yet
- Biofilm Formation and Dispersal in Gram-Positive BacteriaDocument8 pagesBiofilm Formation and Dispersal in Gram-Positive BacteriaRara KhentiNo ratings yet
- The Essential Role of Hydrodynamic Shear Force in The Formation of Biofilm and Granular SludgeDocument13 pagesThe Essential Role of Hydrodynamic Shear Force in The Formation of Biofilm and Granular SludgeRicardo BrandãoNo ratings yet
- Biofilms: Formation & Current Challenges: Group IDocument30 pagesBiofilms: Formation & Current Challenges: Group IAamnah AslamNo ratings yet
- Biofilms in Food IndustryDocument11 pagesBiofilms in Food IndustryKaleem UllahNo ratings yet
- Dependence of Local Cell Density On Concentric RinDocument9 pagesDependence of Local Cell Density On Concentric RinMukhaddam MuhammadNo ratings yet
- Microbial Ecology - Part IDocument8 pagesMicrobial Ecology - Part Isomide kayodeNo ratings yet
- Wilpiszeski et al_2019Document18 pagesWilpiszeski et al_2019scooterneosNo ratings yet
- A Comparative Study of The Nature of Biopolymers Extracted From Anaerobic and Activated SludgesDocument8 pagesA Comparative Study of The Nature of Biopolymers Extracted From Anaerobic and Activated SludgesluizrNo ratings yet
- Examens - Ecotoxicology and MicrobiologyDocument4 pagesExamens - Ecotoxicology and MicrobiologyDMNo ratings yet
- 1 s2.0 S0043135407003193 Main PDFDocument12 pages1 s2.0 S0043135407003193 Main PDFAmira GogoantaNo ratings yet
- Brackets Ceramicos 2Document7 pagesBrackets Ceramicos 2ortodoncia 2018No ratings yet
- Emulsion Separation and Fouling of Electrospun Polyacrylonitrile Membranes For Produced Water Applications - ScienceDirectDocument52 pagesEmulsion Separation and Fouling of Electrospun Polyacrylonitrile Membranes For Produced Water Applications - ScienceDirectMOH AMANNo ratings yet
- Melamine Glyoxylated ResinDocument8 pagesMelamine Glyoxylated ResinRashid Saleem100% (1)
- Project EngineerDocument3 pagesProject Engineerapi-121410515No ratings yet
- Investigation of Graphene/Ag Nanocomposites Synthesis Parameters For Two Different Synthesis MethodsDocument12 pagesInvestigation of Graphene/Ag Nanocomposites Synthesis Parameters For Two Different Synthesis MethodsGera FJNo ratings yet
- Analysis and Conservation of Modern ModelingDocument20 pagesAnalysis and Conservation of Modern ModelingsabuNo ratings yet
- Biodiversity of Cyanobacteria and Green Algae On MonumentsDocument15 pagesBiodiversity of Cyanobacteria and Green Algae On MonumentsWendy PhNo ratings yet
- Scanning Electron Microscopy: A Review and Report of Research in Wood ScienceDocument21 pagesScanning Electron Microscopy: A Review and Report of Research in Wood ScienceRitwik DasNo ratings yet
- Coating 02Document11 pagesCoating 02dhy182No ratings yet
- Project ReportDocument37 pagesProject Reportraghunath reddyparadesNo ratings yet
- Berk-2009-Journal of Forensic SciencesDocument9 pagesBerk-2009-Journal of Forensic SciencesClaudio NovaesNo ratings yet
- Paper 1: AbstractDocument15 pagesPaper 1: AbstractKulsoom KoserNo ratings yet
- Comparative Studies On Corrosion and Tribological Performance of Derakhshandeh2020Document11 pagesComparative Studies On Corrosion and Tribological Performance of Derakhshandeh2020Chercheurs Permanents UrmmNo ratings yet
- Tescan Magna Brochure Uhr SemDocument8 pagesTescan Magna Brochure Uhr SemJulio CasalloNo ratings yet
- The Critical Current Density of Advanced Internal-Mg-Diffusion-Processed Mgb2 WiresDocument9 pagesThe Critical Current Density of Advanced Internal-Mg-Diffusion-Processed Mgb2 WiresprofesorilhamNo ratings yet
- X-Act Brochure PDFDocument2 pagesX-Act Brochure PDFLia Putri TNo ratings yet
- The Effect of Heat Treatment On Interface Properties of S45CDocument8 pagesThe Effect of Heat Treatment On Interface Properties of S45CRamon BrownNo ratings yet
- Ipc2012 90674Document6 pagesIpc2012 90674Marcelo Varejão CasarinNo ratings yet
- Oxford EDS Instruction ManualDocument2 pagesOxford EDS Instruction ManualAnonymous 0tqzNTWyyNo ratings yet
- Polypyrrole-Palladium Systems Prepared in PDCL Aqueous SolutionsDocument10 pagesPolypyrrole-Palladium Systems Prepared in PDCL Aqueous SolutionsemzzNo ratings yet
- CSE007Document159 pagesCSE007arozmanNo ratings yet
- Elektronenmikroskop PDFDocument23 pagesElektronenmikroskop PDFNik EspenNo ratings yet
- Catalyst Support Effects: Gas-Phase Hydrogenation of Phenol Over PalladiumDocument12 pagesCatalyst Support Effects: Gas-Phase Hydrogenation of Phenol Over PalladiumRungrawin NgamkhumNo ratings yet
- 3D Characterization of Porosity and Authigenic Cementation in Triassic Conglomerates-Arenites in The Edvard Grieg Field Using 3D microCT ImagingDocument17 pages3D Characterization of Porosity and Authigenic Cementation in Triassic Conglomerates-Arenites in The Edvard Grieg Field Using 3D microCT ImagingDaniel Mauricio Rojas CaroNo ratings yet
- Corrosion Inhibition at Scribed Locations in Coated AA2024-T3 by Cerium - and DMTD-Loaded Natural Silica Microparticles Under Continuous Immersion and Wet-Dry Cyclic ExposureDocument15 pagesCorrosion Inhibition at Scribed Locations in Coated AA2024-T3 by Cerium - and DMTD-Loaded Natural Silica Microparticles Under Continuous Immersion and Wet-Dry Cyclic ExposureHa Linh VuNo ratings yet
- Laser Scanning MicrosDocument13 pagesLaser Scanning MicrosHà An Đỗ NgọcNo ratings yet
- Tribology InternationalDocument9 pagesTribology InternationalKandice BarrosNo ratings yet
- HTM 08-06 Pathology Labortory Gas SystemsDocument52 pagesHTM 08-06 Pathology Labortory Gas SystemsOmar ZabshNo ratings yet
- Focused Ion BeamDocument90 pagesFocused Ion BeamLotus DarkNo ratings yet
- Kavayitri Bahinabai Chaudhari North Maharashtra University, JalgaonDocument9 pagesKavayitri Bahinabai Chaudhari North Maharashtra University, Jalgaondipakchavan17No ratings yet























































































Download as pdf or txt
You might also like
- Biofilms in Treatment of Waste Water 2Document15 pagesBiofilms in Treatment of Waste Water 2anishapradhan.studNo ratings yet
- Engineering Biofilm Formation and Dispersal: Thomas K. Wood, Seok Hoon Hong and Qun MaDocument8 pagesEngineering Biofilm Formation and Dispersal: Thomas K. Wood, Seok Hoon Hong and Qun MaShampa SenNo ratings yet
- Kishen - Haapasalo - 2010 - Biofilm Models and Methods of Biofilm AssessmentDocument21 pagesKishen - Haapasalo - 2010 - Biofilm Models and Methods of Biofilm AssessmentdrchanyccNo ratings yet
- Corrosion Mic &mitigationDocument6 pagesCorrosion Mic &mitigationprakashNo ratings yet
- Biofilm Models and Methods ofDocument21 pagesBiofilm Models and Methods ofVerghese George MNo ratings yet
- 2010 79 711 FEOFILOVA Fungal Cell Wall MicrobiolDocument10 pages2010 79 711 FEOFILOVA Fungal Cell Wall MicrobiolGebrehiwot GebremedhinNo ratings yet
- MCB 150-C Group 6Document31 pagesMCB 150-C Group 6Angela Kristel DalaoNo ratings yet
- Https Webmail - Med.auth 1Document10 pagesHttps Webmail - Med.auth 1DD DNo ratings yet
- Microcolonies StructureDocument12 pagesMicrocolonies StructuremelanypaguayNo ratings yet
- Fleming and Wingender Biofilm Matrix Nature ReviewsDocument11 pagesFleming and Wingender Biofilm Matrix Nature Reviewsgkhan3No ratings yet
- Vol. 6, Issue 1, January 2018, PharmaTutor, Paper-2Document7 pagesVol. 6, Issue 1, January 2018, PharmaTutor, Paper-2mmmmmNo ratings yet
- Plasma Activated WaterDocument12 pagesPlasma Activated WaterconsendoNo ratings yet
- Biofilms in Endodontic InfectionsDocument10 pagesBiofilms in Endodontic InfectionsValderrama Cisneros José LuisNo ratings yet
- Biofilms and Their Advantages/Disadvantages in Food IndustryDocument7 pagesBiofilms and Their Advantages/Disadvantages in Food IndustryUmarNo ratings yet
- Biodegradacion de XenobioticosDocument8 pagesBiodegradacion de XenobioticosDoc Efra RamirezNo ratings yet
- Biological Fixed Film: Md. Shahinoor Islam, Geelsu Hwang, Yang LiuDocument33 pagesBiological Fixed Film: Md. Shahinoor Islam, Geelsu Hwang, Yang Liusong1088No ratings yet
- Bharti2017 Phototrophic BiofilmsDocument16 pagesBharti2017 Phototrophic BiofilmsSacra PsyntergiaNo ratings yet
- BIOFILMDocument79 pagesBIOFILMrkc456No ratings yet
- Reviews: Bacterial Biofilms: From The Natural Environment To Infectious DiseasesDocument15 pagesReviews: Bacterial Biofilms: From The Natural Environment To Infectious DiseasesJordy KunNo ratings yet
- Sujit - Cife - Role of Biofilm in Aquaculture - FinalDocument55 pagesSujit - Cife - Role of Biofilm in Aquaculture - FinalsushanfcriNo ratings yet
- Biofilm SemDocument64 pagesBiofilm SemmaltiNo ratings yet
- 250 Flemming Wingender ThebiofilmmatrixDocument12 pages250 Flemming Wingender ThebiofilmmatrixdeviimaNo ratings yet
- Chapter 2. Biotechnology Principles of Solid State FermentationDocument52 pagesChapter 2. Biotechnology Principles of Solid State Fermentationm.alejandracruzsalazarNo ratings yet
- InTech-Scanning Electron Microscopy Sem and Environmental Sem Suitable Tools For Study of Adhesion Stage and Biofilm FormationDocument15 pagesInTech-Scanning Electron Microscopy Sem and Environmental Sem Suitable Tools For Study of Adhesion Stage and Biofilm FormationJose Manuel Oliveira PereiraNo ratings yet
- Galley Proof Ms Ijbpas 2021 5610Document9 pagesGalley Proof Ms Ijbpas 2021 5610saisree malladiNo ratings yet
- Biofilm FormationDocument12 pagesBiofilm FormationFhaliesha ZakurNo ratings yet
- Biofilms: Microbial Life On Surfaces: Rodney M. DonlanDocument10 pagesBiofilms: Microbial Life On Surfaces: Rodney M. DonlanKapil JhajhariaNo ratings yet
- Maslowski 2019 Metabolism at The Centre Clinical and Experimental ImmunologyDocument13 pagesMaslowski 2019 Metabolism at The Centre Clinical and Experimental ImmunologyMarcelo MurilloNo ratings yet
- The Soil Microbiome - From Metagenomics To Metaphenomics: SciencedirectDocument7 pagesThe Soil Microbiome - From Metagenomics To Metaphenomics: SciencedirectMiroz RexNo ratings yet
- The Superficial Life of MicrobesDocument3 pagesThe Superficial Life of MicrobesHarveenkaur MalhanNo ratings yet
- Articulo Grupo1Document17 pagesArticulo Grupo1PAULA ANDREA QUICENO GIRALDONo ratings yet
- Bio Film: Emilia Oleandro, Romina Rega, Martina Mugnano, Filomena Nazzaro, Pietro Ferraro, Simonetta GrilliDocument10 pagesBio Film: Emilia Oleandro, Romina Rega, Martina Mugnano, Filomena Nazzaro, Pietro Ferraro, Simonetta GrilliPFENo ratings yet
- Biofilm ReportDocument50 pagesBiofilm ReportMary Joy DalisayNo ratings yet
- Detection of pathogenesis Related proteins- chitinase and β-1,3-glucanase in induced chickpeaDocument4 pagesDetection of pathogenesis Related proteins- chitinase and β-1,3-glucanase in induced chickpeaImmu JiNo ratings yet
- Vanysacker-2013-Microbial Adhesion and BiofilmDocument12 pagesVanysacker-2013-Microbial Adhesion and BiofilmEduard-Marius LungulescuNo ratings yet
- Fmicb 12 675843Document15 pagesFmicb 12 675843Rin ChanNo ratings yet
- Microbiology and Biotechnology ScriptDocument18 pagesMicrobiology and Biotechnology ScriptDavid RăscuţoiNo ratings yet
- CHEMOSTATDocument12 pagesCHEMOSTATHass AnNo ratings yet
- Surface ChemistryDocument15 pagesSurface ChemistryHajime HikariNo ratings yet
- Why Do Microbes Make MineralsDocument40 pagesWhy Do Microbes Make MineralsVelraj ParthibanNo ratings yet
- Tissue Engineering 4Document20 pagesTissue Engineering 4Vidit NarayanNo ratings yet
- Nghien CuuDocument9 pagesNghien CuuNguyen GiaoNo ratings yet
- Biofilm Dispersion: Kendra P. Rumbaugh and Karin SauerDocument16 pagesBiofilm Dispersion: Kendra P. Rumbaugh and Karin SauerTallesNo ratings yet
- Hamedi 2017Document22 pagesHamedi 2017joaovictorandre0609No ratings yet
- Content 9080Document7 pagesContent 9080Andreea MicuNo ratings yet
- Review AdhesionDocument16 pagesReview AdhesionMirella DannaNo ratings yet
- Bio FilmDocument7 pagesBio FilmFREIMUZICNo ratings yet
- Bertrand Et Al. 2021 - Estimation of Pore Dimensions in Lipid Membranes Induced by Peptides and Other Biomolecules A ReviewDocument18 pagesBertrand Et Al. 2021 - Estimation of Pore Dimensions in Lipid Membranes Induced by Peptides and Other Biomolecules A ReviewAmadorRevillaNo ratings yet
- Content ServerDocument10 pagesContent ServerAdit VekariaNo ratings yet
- Biofilm Formation and Dispersal in Gram-Positive BacteriaDocument8 pagesBiofilm Formation and Dispersal in Gram-Positive BacteriaRara KhentiNo ratings yet
- The Essential Role of Hydrodynamic Shear Force in The Formation of Biofilm and Granular SludgeDocument13 pagesThe Essential Role of Hydrodynamic Shear Force in The Formation of Biofilm and Granular SludgeRicardo BrandãoNo ratings yet
- Biofilms: Formation & Current Challenges: Group IDocument30 pagesBiofilms: Formation & Current Challenges: Group IAamnah AslamNo ratings yet
- Biofilms in Food IndustryDocument11 pagesBiofilms in Food IndustryKaleem UllahNo ratings yet
- Dependence of Local Cell Density On Concentric RinDocument9 pagesDependence of Local Cell Density On Concentric RinMukhaddam MuhammadNo ratings yet
- Microbial Ecology - Part IDocument8 pagesMicrobial Ecology - Part Isomide kayodeNo ratings yet
- Wilpiszeski et al_2019Document18 pagesWilpiszeski et al_2019scooterneosNo ratings yet
- A Comparative Study of The Nature of Biopolymers Extracted From Anaerobic and Activated SludgesDocument8 pagesA Comparative Study of The Nature of Biopolymers Extracted From Anaerobic and Activated SludgesluizrNo ratings yet
- Examens - Ecotoxicology and MicrobiologyDocument4 pagesExamens - Ecotoxicology and MicrobiologyDMNo ratings yet
- 1 s2.0 S0043135407003193 Main PDFDocument12 pages1 s2.0 S0043135407003193 Main PDFAmira GogoantaNo ratings yet
- Brackets Ceramicos 2Document7 pagesBrackets Ceramicos 2ortodoncia 2018No ratings yet
- Emulsion Separation and Fouling of Electrospun Polyacrylonitrile Membranes For Produced Water Applications - ScienceDirectDocument52 pagesEmulsion Separation and Fouling of Electrospun Polyacrylonitrile Membranes For Produced Water Applications - ScienceDirectMOH AMANNo ratings yet
- Melamine Glyoxylated ResinDocument8 pagesMelamine Glyoxylated ResinRashid Saleem100% (1)
- Project EngineerDocument3 pagesProject Engineerapi-121410515No ratings yet
- Investigation of Graphene/Ag Nanocomposites Synthesis Parameters For Two Different Synthesis MethodsDocument12 pagesInvestigation of Graphene/Ag Nanocomposites Synthesis Parameters For Two Different Synthesis MethodsGera FJNo ratings yet
- Analysis and Conservation of Modern ModelingDocument20 pagesAnalysis and Conservation of Modern ModelingsabuNo ratings yet
- Biodiversity of Cyanobacteria and Green Algae On MonumentsDocument15 pagesBiodiversity of Cyanobacteria and Green Algae On MonumentsWendy PhNo ratings yet
- Scanning Electron Microscopy: A Review and Report of Research in Wood ScienceDocument21 pagesScanning Electron Microscopy: A Review and Report of Research in Wood ScienceRitwik DasNo ratings yet
- Coating 02Document11 pagesCoating 02dhy182No ratings yet
- Project ReportDocument37 pagesProject Reportraghunath reddyparadesNo ratings yet
- Berk-2009-Journal of Forensic SciencesDocument9 pagesBerk-2009-Journal of Forensic SciencesClaudio NovaesNo ratings yet
- Paper 1: AbstractDocument15 pagesPaper 1: AbstractKulsoom KoserNo ratings yet
- Comparative Studies On Corrosion and Tribological Performance of Derakhshandeh2020Document11 pagesComparative Studies On Corrosion and Tribological Performance of Derakhshandeh2020Chercheurs Permanents UrmmNo ratings yet
- Tescan Magna Brochure Uhr SemDocument8 pagesTescan Magna Brochure Uhr SemJulio CasalloNo ratings yet
- The Critical Current Density of Advanced Internal-Mg-Diffusion-Processed Mgb2 WiresDocument9 pagesThe Critical Current Density of Advanced Internal-Mg-Diffusion-Processed Mgb2 WiresprofesorilhamNo ratings yet
- X-Act Brochure PDFDocument2 pagesX-Act Brochure PDFLia Putri TNo ratings yet
- The Effect of Heat Treatment On Interface Properties of S45CDocument8 pagesThe Effect of Heat Treatment On Interface Properties of S45CRamon BrownNo ratings yet
- Ipc2012 90674Document6 pagesIpc2012 90674Marcelo Varejão CasarinNo ratings yet
- Oxford EDS Instruction ManualDocument2 pagesOxford EDS Instruction ManualAnonymous 0tqzNTWyyNo ratings yet
- Polypyrrole-Palladium Systems Prepared in PDCL Aqueous SolutionsDocument10 pagesPolypyrrole-Palladium Systems Prepared in PDCL Aqueous SolutionsemzzNo ratings yet
- CSE007Document159 pagesCSE007arozmanNo ratings yet
- Elektronenmikroskop PDFDocument23 pagesElektronenmikroskop PDFNik EspenNo ratings yet
- Catalyst Support Effects: Gas-Phase Hydrogenation of Phenol Over PalladiumDocument12 pagesCatalyst Support Effects: Gas-Phase Hydrogenation of Phenol Over PalladiumRungrawin NgamkhumNo ratings yet
- 3D Characterization of Porosity and Authigenic Cementation in Triassic Conglomerates-Arenites in The Edvard Grieg Field Using 3D microCT ImagingDocument17 pages3D Characterization of Porosity and Authigenic Cementation in Triassic Conglomerates-Arenites in The Edvard Grieg Field Using 3D microCT ImagingDaniel Mauricio Rojas CaroNo ratings yet
- Corrosion Inhibition at Scribed Locations in Coated AA2024-T3 by Cerium - and DMTD-Loaded Natural Silica Microparticles Under Continuous Immersion and Wet-Dry Cyclic ExposureDocument15 pagesCorrosion Inhibition at Scribed Locations in Coated AA2024-T3 by Cerium - and DMTD-Loaded Natural Silica Microparticles Under Continuous Immersion and Wet-Dry Cyclic ExposureHa Linh VuNo ratings yet
- Laser Scanning MicrosDocument13 pagesLaser Scanning MicrosHà An Đỗ NgọcNo ratings yet
- Tribology InternationalDocument9 pagesTribology InternationalKandice BarrosNo ratings yet
- HTM 08-06 Pathology Labortory Gas SystemsDocument52 pagesHTM 08-06 Pathology Labortory Gas SystemsOmar ZabshNo ratings yet
- Focused Ion BeamDocument90 pagesFocused Ion BeamLotus DarkNo ratings yet
- Kavayitri Bahinabai Chaudhari North Maharashtra University, JalgaonDocument9 pagesKavayitri Bahinabai Chaudhari North Maharashtra University, Jalgaondipakchavan17No ratings yet