Download as pdf or txt
You might also like
- Sydney Basin Visual Pilot GuideDocument0 pagesSydney Basin Visual Pilot Guidephilridley2No ratings yet
- Detection of Feline Panleukopenia With Antigen Test KitDocument2 pagesDetection of Feline Panleukopenia With Antigen Test KitMuhammad ihwanul usliminNo ratings yet
- Fish and Shell Fish Immunology: Vipin Kumar Verma, Kumari Vandana Rani, Shiva Raj Kumar, Om PrakashDocument9 pagesFish and Shell Fish Immunology: Vipin Kumar Verma, Kumari Vandana Rani, Shiva Raj Kumar, Om PrakashAsmaa MufiidaNo ratings yet
- 143E265B2Document6 pages143E265B2Omobola AmooNo ratings yet
- Jesus Patogenicidad PecesDocument12 pagesJesus Patogenicidad PecesCAMI ARMYNo ratings yet
- Article 1 2222 enDocument14 pagesArticle 1 2222 enIlincaNo ratings yet
- Hal 2020Document6 pagesHal 2020Luis Angel Gonzalez CallirgosNo ratings yet
- Acanthophora SpiciferaDocument8 pagesAcanthophora SpiciferaMuthya BrilianNo ratings yet
- Fish and Shell Fish Immunology: Full Length ArticleDocument9 pagesFish and Shell Fish Immunology: Full Length ArticleLarissa SoaresNo ratings yet
- Antibiotic Susceptibility and Molecular Characterization of Aeromonas Hydrophila From Grass CarpDocument6 pagesAntibiotic Susceptibility and Molecular Characterization of Aeromonas Hydrophila From Grass CarpMiriam LeiNo ratings yet
- Histopathology and Transcriptome Profiling Reveal Features of Immune Responses in Gills and Intestine Induced by Spring Viremia of Carp VirusDocument45 pagesHistopathology and Transcriptome Profiling Reveal Features of Immune Responses in Gills and Intestine Induced by Spring Viremia of Carp VirusJovanaNo ratings yet
- Abalone PDFDocument10 pagesAbalone PDFJose SilvaNo ratings yet
- Qiting Liang, Et Al (2022) Application of Potential Probiotic Strain Streptomyces Sp. SH5 On Anti - Aeromonas Infection in Zebrafish LarvaeDocument11 pagesQiting Liang, Et Al (2022) Application of Potential Probiotic Strain Streptomyces Sp. SH5 On Anti - Aeromonas Infection in Zebrafish LarvaeAldy KiatNo ratings yet
- Zhang 2012Document13 pagesZhang 2012aji gumelarNo ratings yet
- Shrimp ImmunostimulantsDocument27 pagesShrimp ImmunostimulantsSyed Omer AliNo ratings yet
- Cristinalopesdocarmo 2020Document7 pagesCristinalopesdocarmo 2020Leandro DouglasNo ratings yet
- 10.1007@s10123 019 00075 3Document12 pages10.1007@s10123 019 00075 3Sandy Wahyu DNo ratings yet
- ImuDocument28 pagesImuLukmanAnugrahNo ratings yet
- Uso de Vacunas de ADN vs. Salmonella EntericaDocument10 pagesUso de Vacunas de ADN vs. Salmonella EntericaalferezhassanNo ratings yet
- PotatoBiotec 2009Document6 pagesPotatoBiotec 2009Erika WolskiNo ratings yet
- Anshour Et AlDocument12 pagesAnshour Et AlAdam PratamaNo ratings yet
- Li 2012Document7 pagesLi 2012Yudho AdhitomoNo ratings yet
- Massive Mortality Associated With StreptDocument8 pagesMassive Mortality Associated With StreptAbir BoukhennouNo ratings yet
- Fish & Shell Fish Immunology: Full Length ArticleDocument12 pagesFish & Shell Fish Immunology: Full Length ArticleatikamarisaNo ratings yet
- TMP B7 EBDocument9 pagesTMP B7 EBFrontiersNo ratings yet
- Fish & Shell Fish ImmunologyDocument7 pagesFish & Shell Fish ImmunologyBaskaran ViswanathanNo ratings yet
- Martínez Et Al 2018Document10 pagesMartínez Et Al 2018Adrian Melgratti JobsonNo ratings yet
- ParasiteDocument5 pagesParasiteDeLiza SuhaaNo ratings yet
- Enrichment of Common Carp (Cyprinus Carpio) Fingerlings Diet With Psidium GuajavaDocument29 pagesEnrichment of Common Carp (Cyprinus Carpio) Fingerlings Diet With Psidium GuajavaCindy Guliana Hernandez ArguelloNo ratings yet
- Angulo Et Al., 2020-LDocument11 pagesAngulo Et Al., 2020-LLuis Alberto Canul KuNo ratings yet
- LSHTM Research Online: Usage GuidelinesDocument7 pagesLSHTM Research Online: Usage GuidelinesIgor BaltaNo ratings yet
- The Protective Effect of Oral Specific Yolk Antibody On Mandarin FishDocument18 pagesThe Protective Effect of Oral Specific Yolk Antibody On Mandarin FishJovanaNo ratings yet
- De Sá Hyacienth Et Al. - 2020 - Endopleura Uchi (Huber) Cuatrec. A Medicinal Plant For Gynecological Treatments - A Reproductive ToxicitDocument12 pagesDe Sá Hyacienth Et Al. - 2020 - Endopleura Uchi (Huber) Cuatrec. A Medicinal Plant For Gynecological Treatments - A Reproductive ToxicitSALAZAR JANSEMNo ratings yet
- Development of Attenuated Erythromycin Resistant Streptococcus Agalactiae Vaccine For Tilapia (Oreochromis Niloticus) CultureDocument9 pagesDevelopment of Attenuated Erythromycin Resistant Streptococcus Agalactiae Vaccine For Tilapia (Oreochromis Niloticus) CultureSTRUGGLE KEEPNo ratings yet
- Shivaraj2012 PDFDocument1 pageShivaraj2012 PDFffNo ratings yet
- Emecheta 20181Document9 pagesEmecheta 20181Ahmed HamdyNo ratings yet
- s12639 020 01217 5Document4 pagess12639 020 01217 5Iryna ZapekaNo ratings yet
- The Antioxidant, Immunomodulatory, and Anti Inflammatory Activities of Spirulina: An OverviewDocument24 pagesThe Antioxidant, Immunomodulatory, and Anti Inflammatory Activities of Spirulina: An OverviewPlacido Rojas FrancoNo ratings yet
- Mco 12 06 0533Document8 pagesMco 12 06 0533Anonymous Wcj4C3jNo ratings yet
- Sciadv Aba2578Document10 pagesSciadv Aba2578Pratiwi ZikriNo ratings yet
- Anaplasma Bovis (Ehrlichia Bovis) Infection in A Buffalo-A Case ReportDocument4 pagesAnaplasma Bovis (Ehrlichia Bovis) Infection in A Buffalo-A Case ReportshivarajuNo ratings yet
- Articulo GabrielaDocument10 pagesArticulo GabrielaGaby MoralesNo ratings yet
- TMP D351Document5 pagesTMP D351FrontiersNo ratings yet
- 2022 The Combined Use of PediococcusDocument10 pages2022 The Combined Use of Pediococcuscleon79No ratings yet
- Fcimb 11 643312Document13 pagesFcimb 11 643312Edson EscamillaNo ratings yet
- Cryptosporidiosis in Ruminants: Update and Current Therapeutic ApproachesDocument8 pagesCryptosporidiosis in Ruminants: Update and Current Therapeutic ApproachesDrivailaNo ratings yet
- Effect of A Potential Probiotic Candidate Enterococcus Faecalis-1 On Growth Performance, Intestinal Microbiota, and Immune Response of Commercial Broiler ChickensDocument10 pagesEffect of A Potential Probiotic Candidate Enterococcus Faecalis-1 On Growth Performance, Intestinal Microbiota, and Immune Response of Commercial Broiler ChickenszawarftNo ratings yet
- DocumentDocument12 pagesDocumentariefNo ratings yet
- Developmental and Comparative ImmunologyDocument13 pagesDevelopmental and Comparative ImmunologyMuchammadIbrahimUlilAbshorNo ratings yet
- Expression of The Recombinant Enterocin E-760 in Pichia Pastoris X33 and Its Antimicrobial Activity Towards Listeria MonocytogenesDocument10 pagesExpression of The Recombinant Enterocin E-760 in Pichia Pastoris X33 and Its Antimicrobial Activity Towards Listeria MonocytogenesSri FamilasariNo ratings yet
- Survivability of Low Pathogenic (H9N2) Avian Influenza Virus in Water in The Presence of Atyopsis Moluccensis (Bamboo Shrimp)Document6 pagesSurvivability of Low Pathogenic (H9N2) Avian Influenza Virus in Water in The Presence of Atyopsis Moluccensis (Bamboo Shrimp)Johanne May Capitulo LopezNo ratings yet
- 14 Pathogenicity of Aeromonas Hydrophila To Blunt SnoutDocument7 pages14 Pathogenicity of Aeromonas Hydrophila To Blunt SnoutRafi Adi SNo ratings yet
- Anti-Influenza Activity of Elderberry - Sambucus NigraDocument8 pagesAnti-Influenza Activity of Elderberry - Sambucus NigraDiego RodriguezNo ratings yet
- InulinaDocument6 pagesInulinacleon79No ratings yet
- Amalraj Jeyaprabha: Hunan Daxue Xuebao/Journal of Hunan University Natural SciencesDocument20 pagesAmalraj Jeyaprabha: Hunan Daxue Xuebao/Journal of Hunan University Natural Sciencesindex PubNo ratings yet
- Fish & Shellfish ImmunologyDocument8 pagesFish & Shellfish ImmunologyFebby da costaNo ratings yet
- Chinese Herbs (Astragalus Membranaceus and Lonicera Japonica) and Boron Enhance The Non-Specific Immune Response of Nile Tilapia (Oreochromis Niloticus) and Resistance Against Aeromonas HydrophilaDocument9 pagesChinese Herbs (Astragalus Membranaceus and Lonicera Japonica) and Boron Enhance The Non-Specific Immune Response of Nile Tilapia (Oreochromis Niloticus) and Resistance Against Aeromonas HydrophilaCecilia Almazan SanchezNo ratings yet
- Chinese Herbs (Astragalus Radix and Ganoderma Lucidum) Enhance Immune Response of Carp, Cyprinus Carpio, and Protection Against Aeromonas Hydrophila. Fish Shellfish ImmunolDocument7 pagesChinese Herbs (Astragalus Radix and Ganoderma Lucidum) Enhance Immune Response of Carp, Cyprinus Carpio, and Protection Against Aeromonas Hydrophila. Fish Shellfish ImmunolCecilia Almazan SanchezNo ratings yet
- Author's Accepted Manuscript: FicusDocument33 pagesAuthor's Accepted Manuscript: FicusMaria DudauNo ratings yet
- The Biology of Fungi Impacting Human Health: A Tropical Asia-Pacific PerspectiveFrom EverandThe Biology of Fungi Impacting Human Health: A Tropical Asia-Pacific PerspectiveRating: 5 out of 5 stars5/5 (1)
- All India Aakash Test Series For NEET - 2021 TEST - 6 (Code-C)Document32 pagesAll India Aakash Test Series For NEET - 2021 TEST - 6 (Code-C)Kavyatharsheni S XI-B 46No ratings yet
- Crafting and Executing Strategy 19th Edition Thompson Test Bank 1Document193 pagesCrafting and Executing Strategy 19th Edition Thompson Test Bank 1raymond100% (43)
- Throw Training ProgramDocument3 pagesThrow Training ProgramSir Manny100% (1)
- Unit 2 RevisiónDocument6 pagesUnit 2 Revisióndayana Carolina sanchez arauzNo ratings yet
- Caterpillar Xqe 20 Kva: Powered by ExcellenceDocument2 pagesCaterpillar Xqe 20 Kva: Powered by ExcellenceMiu MiuNo ratings yet
- Capt - Pradeep Correa - Growth of The India Economy and Ship BDocument33 pagesCapt - Pradeep Correa - Growth of The India Economy and Ship BacoopsNo ratings yet
- OK-LN SeriesDocument10 pagesOK-LN SeriesSuresh GoddetiNo ratings yet
- The Bad Food StoryDocument3 pagesThe Bad Food StoryraamsNo ratings yet
- Material and DesignDocument2 pagesMaterial and DesignrafadannNo ratings yet
- Cajetin Word VacioDocument7 pagesCajetin Word VaciogolfersantosNo ratings yet
- Peta CalculusDocument2 pagesPeta CalculusAbby NavarroNo ratings yet
- Top Companies of IndiaDocument19 pagesTop Companies of IndiaNidhi_Rawat_7890100% (1)
- Pub - Logarithmic Forms and Diophantine Geometry New Mat PDFDocument210 pagesPub - Logarithmic Forms and Diophantine Geometry New Mat PDFgopanov100% (2)
- Rittal Technical System Catalogue Ri4Power 5 2020Document92 pagesRittal Technical System Catalogue Ri4Power 5 2020Zivorad SerafimoskiNo ratings yet
- Static Electricity Lesson PlanDocument14 pagesStatic Electricity Lesson PlanKarnadi WibowoNo ratings yet
- Acceleration WorksheetDocument3 pagesAcceleration WorksheetJulia RibeiroNo ratings yet
- CH 4.2 - Mass BrainGuard IMAM-12Document21 pagesCH 4.2 - Mass BrainGuard IMAM-12tolerancelost607100% (1)
- RBDG MAN 027 0105 - Environment - RedactedDocument23 pagesRBDG MAN 027 0105 - Environment - RedactedSezgin İçaNo ratings yet
- The Hour I First Believed - Slate Star CodexDocument37 pagesThe Hour I First Believed - Slate Star CodexSunshine FrankensteinNo ratings yet
- Mikyas. 2020. Positive Science or Interpretive UnderstandingDocument16 pagesMikyas. 2020. Positive Science or Interpretive UnderstandingMikyas AberaNo ratings yet
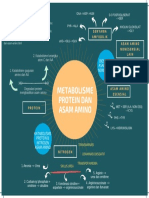
- Metabolisme Protein Dan Asam Amino: Mind Map TM Ii/Idk IDocument1 pageMetabolisme Protein Dan Asam Amino: Mind Map TM Ii/Idk ISania qurrataNo ratings yet
- Lista de Medicamentos Esenciales para Emergencias Radiológicas y NuclearesDocument66 pagesLista de Medicamentos Esenciales para Emergencias Radiológicas y NuclearesMartin GonzalezNo ratings yet
- Pemanfaatan Lahan Pekarangan Dengan Budidaya Tanaman Buah Dalam Pot (Tabulampot) Di Desa Gesik, Kecamatan Tengah Tani, Kabupaten CirebonDocument6 pagesPemanfaatan Lahan Pekarangan Dengan Budidaya Tanaman Buah Dalam Pot (Tabulampot) Di Desa Gesik, Kecamatan Tengah Tani, Kabupaten CirebonUmi TrisnaningsihNo ratings yet
- NepalDocument22 pagesNepalDarya KhanNo ratings yet
- September 4, 2019: Vladimir S. MiraranDocument16 pagesSeptember 4, 2019: Vladimir S. MiraranPinkz Trinidad TalionNo ratings yet
- Concreep 10 1Document48 pagesConcreep 10 1NachoNo ratings yet
- Kenwood B62-2299-20Document2 pagesKenwood B62-2299-20King MaxNo ratings yet
- Countable and Uncountable NounsDocument7 pagesCountable and Uncountable NounsValentine NNo ratings yet
- Bridges & Structures: Technical Manual For Design and Construction of Road Tunnels - Civil ElementsDocument12 pagesBridges & Structures: Technical Manual For Design and Construction of Road Tunnels - Civil ElementsSambhav PoddarNo ratings yet