Download as pdf or txt
You might also like
- ملف 58 معدلDocument58 pagesملف 58 معدلraghdah alhogailNo ratings yet
- Drosophila Lab ReportDocument7 pagesDrosophila Lab ReportTim HepburnNo ratings yet
- Mitosis in The Root Meristem of Onion, Allium CepaDocument10 pagesMitosis in The Root Meristem of Onion, Allium CepaJoshuaNo ratings yet
- 10 1006@excr 1996 0098Document10 pages10 1006@excr 1996 0098ivoflorentino8810No ratings yet
- Kinesin Follows The Microtubule's Protofilament Axis: Meyh6fer, Ronald A. Milligan, HowardDocument11 pagesKinesin Follows The Microtubule's Protofilament Axis: Meyh6fer, Ronald A. Milligan, HowardAlessandro BianchessiNo ratings yet
- Mitochondrial Energy Metabolism in Baby Hamster Kidney (BHK-21/C13) Cells Treated With Karnozin EXTRA®Document7 pagesMitochondrial Energy Metabolism in Baby Hamster Kidney (BHK-21/C13) Cells Treated With Karnozin EXTRA®Garbuz ElenaNo ratings yet
- 331 FullDocument10 pages331 FullJean Pierre Chastre LuzaNo ratings yet
- Carbon Nanotubes As Intracellular Protein Transporters: Generality and Biological FunctionalityDocument6 pagesCarbon Nanotubes As Intracellular Protein Transporters: Generality and Biological Functionality227986No ratings yet
- Cell CycleDocument10 pagesCell Cyclesatheeshpharma6No ratings yet
- Different Types of Cell-to-Cell Connections Mediated by Nanotubular StructuresDocument10 pagesDifferent Types of Cell-to-Cell Connections Mediated by Nanotubular StructuresHans SprungfeldNo ratings yet
- Myosin Motors Drive Long Range AlignmentDocument12 pagesMyosin Motors Drive Long Range AlignmentAlexNo ratings yet
- J Cell Biol 1994 Rieder 1301 10Document10 pagesJ Cell Biol 1994 Rieder 1301 10TarunSreerajNo ratings yet
- Cito Esqueleto MeiosisDocument10 pagesCito Esqueleto MeiosisYair Rodriguez SantiagoNo ratings yet
- BZ Lab 3Document11 pagesBZ Lab 3Alexa Jean D. HonrejasNo ratings yet
- In Vitro Mesenchymal Stem Cell Differentiation After Mechanical StimulationDocument10 pagesIn Vitro Mesenchymal Stem Cell Differentiation After Mechanical StimulationBastion ReinhartNo ratings yet
- Jemt 10165Document11 pagesJemt 10165Alana VinhoteNo ratings yet
- J Cell Sci-1993-Perou-99-107Document9 pagesJ Cell Sci-1993-Perou-99-107glodovichiNo ratings yet
- Cell CycleDocument19 pagesCell CycleMoffat KaprezzoNo ratings yet
- Jurnal Inter Mitosis 5Document7 pagesJurnal Inter Mitosis 5Muhammad Choiruddin ZuhriNo ratings yet
- Efficient Method of Cloning The Obligate Intracellular Bacterium Coxiella BurnetiiDocument7 pagesEfficient Method of Cloning The Obligate Intracellular Bacterium Coxiella BurnetiiDiorella Marie López GonzálezNo ratings yet
- Jbacter00584 0499Document14 pagesJbacter00584 0499YO :)No ratings yet
- Vakuola Kontraktil PDFDocument17 pagesVakuola Kontraktil PDFShin Oriflame MalangNo ratings yet
- Cefotiam in VivoDocument10 pagesCefotiam in VivoVirus HepatitisCNo ratings yet
- Nature 1963 197 452Document6 pagesNature 1963 197 452RaffaNo ratings yet
- Conserved Microtubule-Actin Interactions in Cell Movement and Morphogenesis 03Document11 pagesConserved Microtubule-Actin Interactions in Cell Movement and Morphogenesis 03Sonia Barbosa CornelioNo ratings yet
- Theriogenology: Hiroki Kuwayama, Yoshiaki Tanabe, Teruhiko Wakayama, Satoshi KishigamiDocument7 pagesTheriogenology: Hiroki Kuwayama, Yoshiaki Tanabe, Teruhiko Wakayama, Satoshi KishigamiJOAO VITOR DA SILVA VIANANo ratings yet
- Ultrastructure and Cytochemistry of Dormant Basidiospores of Psilocybe Cubensis1Document13 pagesUltrastructure and Cytochemistry of Dormant Basidiospores of Psilocybe Cubensis1Dayon BochniaNo ratings yet
- CUOncology Society Poster A1 PortraitDocument1 pageCUOncology Society Poster A1 Portraitkenli330200No ratings yet
- Cromatina y ApoptosisDocument11 pagesCromatina y ApoptosisOperacion ConocimientoNo ratings yet
- Transformation Dibutyryl: Morphological of Adenosine Cyclic 3':5'-Monophosphate and TestosteroneDocument4 pagesTransformation Dibutyryl: Morphological of Adenosine Cyclic 3':5'-Monophosphate and TestosteroneJean Pierre Chastre LuzaNo ratings yet
- A Few Approaches To The Study of Endomembranes: Insights Gained From AutoradiographyDocument10 pagesA Few Approaches To The Study of Endomembranes: Insights Gained From AutoradiographyDanie MorenoNo ratings yet
- Bacteriasextracelular EvasiontuberculosisrespinmuneDocument9 pagesBacteriasextracelular EvasiontuberculosisrespinmuneMauricio RodríguezNo ratings yet
- JC 672400Document19 pagesJC 672400Bastion ReinhartNo ratings yet
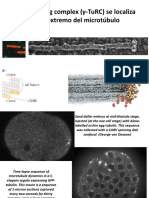
- Γ-Tubulin Ring Complex (Γ-Turc) Se Localiza En Un Extremo Del MicrotúbuloDocument14 pagesΓ-Tubulin Ring Complex (Γ-Turc) Se Localiza En Un Extremo Del MicrotúbuloFELIPE VALBUENANo ratings yet
- Synthesis of Cellulose by Acetobacter XyDocument16 pagesSynthesis of Cellulose by Acetobacter XyKaanNo ratings yet
- 1 s2.0 S0092867418300576 MainextDocument17 pages1 s2.0 S0092867418300576 MainextJOHN ALEJANDRO DIAZ RODRIGUEZNo ratings yet
- Methyl-β-Cyclodextrin Impairs the Monocyte-Adhering Ability of Endothelial Cells by Down-Regulating Adhesion Molecules and Caveolae and Reorganizing the Actin CytoskeletonDocument6 pagesMethyl-β-Cyclodextrin Impairs the Monocyte-Adhering Ability of Endothelial Cells by Down-Regulating Adhesion Molecules and Caveolae and Reorganizing the Actin CytoskeletonGiggly HadidNo ratings yet
- tmpD44A TMPDocument15 pagestmpD44A TMPFrontiersNo ratings yet
- Rat Adult Stem Cells (Marrow Stromal Cells) Engraft and Differentiate in Chick Embryos Without Evidence of Cell FusionDocument4 pagesRat Adult Stem Cells (Marrow Stromal Cells) Engraft and Differentiate in Chick Embryos Without Evidence of Cell Fusion1alfredo12100% (1)
- The Distribution and Behaviour of Intravenously inDocument10 pagesThe Distribution and Behaviour of Intravenously inBruce RoserNo ratings yet
- BFE Unit 5Document26 pagesBFE Unit 5DanielNo ratings yet
- Adaptation To High Rates of Chromosomal Instability and Aneuploidy Through Multiple Pathways in Budding YeastDocument19 pagesAdaptation To High Rates of Chromosomal Instability and Aneuploidy Through Multiple Pathways in Budding YeastFernandoNo ratings yet
- Mechanisms and Functions of Endocytosis: The Journal of Cell Biology February 2008Document6 pagesMechanisms and Functions of Endocytosis: The Journal of Cell Biology February 2008Patrisia AyuNo ratings yet
- Experiment and Mechanism Research of SKOV3 Cancer Cell Apoptosis Induced by Nanosecond Pulsed Electric FieldDocument4 pagesExperiment and Mechanism Research of SKOV3 Cancer Cell Apoptosis Induced by Nanosecond Pulsed Electric Fieldjuliogomez008No ratings yet
- Phototaxis Beyond Turning: Persistent Accumulation and Response Acclimation of The Microalga ChlamydomonasDocument7 pagesPhototaxis Beyond Turning: Persistent Accumulation and Response Acclimation of The Microalga Chlamydomonasalfredc20No ratings yet
- Journal of Histochemistry & Cytochemistry: Reduction of Lipofuscin-Like Autofluorescence in Fluorescently Labeled TissueDocument13 pagesJournal of Histochemistry & Cytochemistry: Reduction of Lipofuscin-Like Autofluorescence in Fluorescently Labeled TissueLovadi Emese IldikóNo ratings yet
- Origin of The Cell Nucleus, Mitosis and Sex Roles of Intracellular CoevolutionDocument78 pagesOrigin of The Cell Nucleus, Mitosis and Sex Roles of Intracellular CoevolutionnolosupeNo ratings yet
- Hum. Reprod.-1996-Wyss-1992-7Document6 pagesHum. Reprod.-1996-Wyss-1992-7Juliana GuerraNo ratings yet
- Cancer Res 1961 Dales 193 7Document9 pagesCancer Res 1961 Dales 193 7documentosdescribdNo ratings yet
- TMP 94 C1Document121 pagesTMP 94 C1FrontiersNo ratings yet
- Formation of Spicules by Sclerocytes From The Fres PDFDocument9 pagesFormation of Spicules by Sclerocytes From The Fres PDFMuhammad Rizqi AdityansyahNo ratings yet
- Prevention of Apoptosis by Bcl-2: Release of Cytochrome C From Mitochondria BlockedDocument5 pagesPrevention of Apoptosis by Bcl-2: Release of Cytochrome C From Mitochondria BlockedKevin MuñozNo ratings yet
- Clonogenic Assay of Cells in Vitro - Nature Protocol 2006Document5 pagesClonogenic Assay of Cells in Vitro - Nature Protocol 2006Thomas SecherNo ratings yet
- Gould y Simanis. Formación Del Septo en LevadurasDocument14 pagesGould y Simanis. Formación Del Septo en LevadurasSaila Viridiana CazaresNo ratings yet
- Cell Cycle Karyotyping Lab ReportDocument11 pagesCell Cycle Karyotyping Lab ReportRichie JustinNo ratings yet
- Cell BiolDocument6 pagesCell BiolHamza AhmedNo ratings yet
- Light The: Regulation of Cell Cycle inDocument7 pagesLight The: Regulation of Cell Cycle inTeresa MataNo ratings yet
- 653 FullDocument17 pages653 Fullmantenimiento.servelshNo ratings yet
- Presentation of Learning in BacteriaDocument21 pagesPresentation of Learning in BacteriaVijay IyerNo ratings yet
- 1 Repiska 2010Document6 pages1 Repiska 2010Thamyres BrancoNo ratings yet
- Su-Jung Chang, Su-Min Tseng, and Hsin-Yiu ChouDocument9 pagesSu-Jung Chang, Su-Min Tseng, and Hsin-Yiu ChouMirza ČelebičićNo ratings yet
- Protein tyrosine phosphatase interacting protein 51 during migration in HaCat cellsFrom EverandProtein tyrosine phosphatase interacting protein 51 during migration in HaCat cellsNo ratings yet
- B430 EliurusphylDocument70 pagesB430 EliurusphylIstvan DanielNo ratings yet
- March 09, 2023 - DNA AND RNA STRUCTURESDocument4 pagesMarch 09, 2023 - DNA AND RNA STRUCTURESJunaisa OrabNo ratings yet
- Timeline QuestionsDocument2 pagesTimeline Questionsvee kims100% (1)
- New Sexual SelectionDocument75 pagesNew Sexual SelectionMala-Maria STAVRESCU-BEDIVANNo ratings yet
- Basic Molecular Biology: Gene StructureDocument36 pagesBasic Molecular Biology: Gene StructureLutfil HadiNo ratings yet
- HMPE 317 (Health Spa and Wellness) TopicDocument2 pagesHMPE 317 (Health Spa and Wellness) TopicGretchen LaurenteNo ratings yet
- Genetically Modified OrganismsDocument2 pagesGenetically Modified OrganismsGeronimo MagparoNo ratings yet
- IgY Proteome Partitioning SolutionsDocument12 pagesIgY Proteome Partitioning SolutionsmfsmarcilioNo ratings yet
- Cellular and Molecular Biology Sample Course ScheduleDocument1 pageCellular and Molecular Biology Sample Course ScheduleSteamaccount aimitoNo ratings yet
- BT Cotton Scenario in India: M.Narayanan 2019508202 GPB 511Document22 pagesBT Cotton Scenario in India: M.Narayanan 2019508202 GPB 511narayananNo ratings yet
- Branches ListDocument3 pagesBranches ListSamantha RullaNo ratings yet
- NEET Biotechnology & It's Application Important QuestionsDocument21 pagesNEET Biotechnology & It's Application Important QuestionsAnimesh KhaNo ratings yet
- Electroporation Method PrincipleDocument28 pagesElectroporation Method PrincipleHai ThanhNo ratings yet
- University of Santo Tomas The Graduate School: An Outcomes-Based Learning ProgramDocument7 pagesUniversity of Santo Tomas The Graduate School: An Outcomes-Based Learning ProgramKathrina Bartido ViescaNo ratings yet
- Protease Revisting The Types and Potential PDFDocument8 pagesProtease Revisting The Types and Potential PDFValeriu MunteanuNo ratings yet
- BINBINON - STS Assignment 3Document4 pagesBINBINON - STS Assignment 3Tom Ivan BinbinonNo ratings yet
- Application of Biology (ZOOLOGY)Document10 pagesApplication of Biology (ZOOLOGY)md dtlNo ratings yet
- Year 5 Plants and Their Life Cycles Guidance SheetDocument1 pageYear 5 Plants and Their Life Cycles Guidance SheetMasthankhan PatanNo ratings yet
- Namets Past Q&aDocument190 pagesNamets Past Q&aZechariah NggitaNo ratings yet
- Induced Mutations and Crop ImprovementDocument26 pagesInduced Mutations and Crop ImprovementPIP AND ZIPNo ratings yet
- Biology: Cambridge International Examinations International General Certificate of Secondary EducationDocument20 pagesBiology: Cambridge International Examinations International General Certificate of Secondary Educationmath magicNo ratings yet
- GenesDocument4 pagesGenesLittle AlligatorNo ratings yet
- Europass Curriculum Vitae: Personal Information José Eduardo Ferreira AraújoDocument5 pagesEuropass Curriculum Vitae: Personal Information José Eduardo Ferreira AraújoARUN PAGIDIMARRINo ratings yet
- Science ReviewerDocument4 pagesScience Reviewerkira xxxNo ratings yet
- Link Belt Wheel Loader l125 Electrical Schematic Hydraulic SchematicDocument22 pagesLink Belt Wheel Loader l125 Electrical Schematic Hydraulic Schematicchelseagomez020399mdi100% (131)
- Activity No. 3 Proteins SasDocument2 pagesActivity No. 3 Proteins SasHecStoryNo ratings yet
- Biochemistry Answer Key-GREEN PACOPDocument18 pagesBiochemistry Answer Key-GREEN PACOPailene agustinNo ratings yet
- Animal Diversity Invertebrates (Zoo201) HandoutsDocument151 pagesAnimal Diversity Invertebrates (Zoo201) HandoutsSufyan khanNo ratings yet