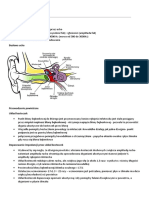
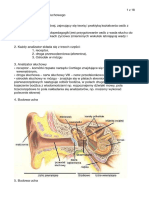
Download as pdf or txt
You might also like
- Fale Stojace W StrunachDocument16 pagesFale Stojace W Strunacha bcNo ratings yet
- Psychoakustyka W PigulceDocument41 pagesPsychoakustyka W PigulcePawel Andryszczyk100% (1)
- Zmysł SłuchuDocument6 pagesZmysł SłuchuKatarzyna Dyba NowotnyNo ratings yet
- Tworzenie GłosuDocument2 pagesTworzenie GłosuMart MalNo ratings yet
- SłuchDocument3 pagesSłuchiza-iza-666No ratings yet
- Lekcja14 AkustykaDocument5 pagesLekcja14 AkustykaKURZA TWARZNo ratings yet
- Fizjologia - Zmysł SłuchuDocument11 pagesFizjologia - Zmysł SłuchuKasia RapaczNo ratings yet
- Narząd Słuchu - FizjologiaDocument4 pagesNarząd Słuchu - FizjologiaGosia SciosNo ratings yet
- Biologia-kl-VII 05 05 (Ucho) PDFDocument4 pagesBiologia-kl-VII 05 05 (Ucho) PDFAnastyshin ФісташкаNo ratings yet
- Ucho - Narzad Słuchu I RównowagiDocument4 pagesUcho - Narzad Słuchu I RównowagiPaulina MatyjaszekNo ratings yet
- Ucho Narzad Sluchu I Rownowagi 2Document19 pagesUcho Narzad Sluchu I Rownowagi 2gaskafilip30No ratings yet
- Fale Dźwiękowe. Na WyrywkiDocument2 pagesFale Dźwiękowe. Na WyrywkiLera WwsNo ratings yet
- SłuchDocument4 pagesSłuchjulia.staszczuk33No ratings yet
- Bio Psycholog I ADocument2 pagesBio Psycholog I Azuzanna bNo ratings yet
- Uklad Oddechowy I Jego FunkcjeDocument21 pagesUklad Oddechowy I Jego FunkcjeEla OnderskaNo ratings yet
- Fale DzwiekoweDocument11 pagesFale Dzwiekowekacperg0608No ratings yet
- Fizjologia Narządu Słuchu I RównowagiDocument31 pagesFizjologia Narządu Słuchu I RównowagiQba888No ratings yet
- UchoDocument8 pagesUchoKrisNo ratings yet
- Čulo SluhaDocument11 pagesČulo SluhaCe CaNo ratings yet
- Kalat - SłuchDocument4 pagesKalat - Słuchanalfab3tyzmNo ratings yet
- O Pracy Głosem Słów KilkaDocument5 pagesO Pracy Głosem Słów KilkaMart MalNo ratings yet
- CzyneuronylubiaciszeDocument14 pagesCzyneuronylubiaciszePatrycja LamczykNo ratings yet
- Wykład 1Document8 pagesWykład 1Wiktoria GawronNo ratings yet
- Świat Pełen DźwiękówDocument15 pagesŚwiat Pełen DźwiękówFareSofaNo ratings yet
- Zmysł Słuchu - NeuroExpert - Encyklopedia NeurofizjologiiDocument1 pageZmysł Słuchu - NeuroExpert - Encyklopedia Neurofizjologiianiab20062006No ratings yet
- Badanie Drżenia GłosowegoDocument1 pageBadanie Drżenia Głosowegojulianna.lisownaNo ratings yet
- Aparat Glosowy - Cwiczymy Glos-1Document27 pagesAparat Glosowy - Cwiczymy Glos-1Monika KrzyżykowskaNo ratings yet
- Fizyka - Drgania I FaleDocument4 pagesFizyka - Drgania I FalelucjakmiecinskaNo ratings yet
- Więzadła Głosowe I Powstawanie DźwiękuDocument2 pagesWięzadła Głosowe I Powstawanie Dźwiękumisiek963963No ratings yet
- Co SłychaćDocument27 pagesCo SłychaćPiotrek WalczakNo ratings yet
- Antaomioa Kolokwium Część 10,11,12,13Document12 pagesAntaomioa Kolokwium Część 10,11,12,13lilyroseNo ratings yet
- Jak OddychamyllDocument11 pagesJak OddychamyllKacper OgonowskiNo ratings yet
- Emisja GłosuDocument6 pagesEmisja Głosumysza_qpaNo ratings yet
- Aparat GłosowyDocument3 pagesAparat GłosowyWeronika PawelecNo ratings yet
- Układ Oddechowy cz.1Document33 pagesUkład Oddechowy cz.1wiktoriazyzykNo ratings yet
- Roz 2Document34 pagesRoz 2Sandra TurczyńskaNo ratings yet
- Logopedia - Silesiana r2012 t1 s104 110Document8 pagesLogopedia - Silesiana r2012 t1 s104 110Głos - źródło poznaniaNo ratings yet
- EMISJA GŁOSU - M. JUREWICZ - KsiążkaDocument55 pagesEMISJA GŁOSU - M. JUREWICZ - KsiążkaWeronika PawelecNo ratings yet
- Robert Harrak - Głośniki - PodstawyDocument23 pagesRobert Harrak - Głośniki - PodstawymasterpogoNo ratings yet
- NotatkiDocument19 pagesNotatkiantypatiaNo ratings yet
- 09 Danel 68-75+Document8 pages09 Danel 68-75+karczakmichalNo ratings yet
- 4317622ee263da7045df6fe616edac87Document1 page4317622ee263da7045df6fe616edac87Jan ZakrzewskiNo ratings yet
- Drgania I FaleDocument5 pagesDrgania I FaleMartyna KomorniczakNo ratings yet
- FiO NR 10Document17 pagesFiO NR 10balcerakjerzNo ratings yet
- Słownik UsgDocument103 pagesSłownik Usgzajeciowarutyna20No ratings yet
- Prezentacja Fizjologia OddychaniaDocument14 pagesPrezentacja Fizjologia OddychaniaAndrzejNo ratings yet
- UchoDocument1 pageUchoAntek KotarskiNo ratings yet
- Układ OdddechowyDocument39 pagesUkład OdddechowyZuzanna KasprzakNo ratings yet
- Układ Oddechowy 1Document65 pagesUkład Oddechowy 1AndrzejNo ratings yet
- Historia Muzyki Semestr IDocument12 pagesHistoria Muzyki Semestr IPeterNo ratings yet
- Ustnik Oddech ZadecieDocument2 pagesUstnik Oddech ZadecieGrzegorz OlewińskiNo ratings yet
- Bluesowa Harmonijka UstnaDocument19 pagesBluesowa Harmonijka UstnaStyrmir NatrakhNo ratings yet
- SURDOPEDAGOGIKADocument18 pagesSURDOPEDAGOGIKAKarolina SadowskaNo ratings yet
- Akustyka OdpDocument11 pagesAkustyka Odpprdaria1mportantNo ratings yet
- Wykład 1 - Emisja GłosuDocument3 pagesWykład 1 - Emisja Głosuszymonjania765No ratings yet
- Jak Powstaje GlosDocument20 pagesJak Powstaje GlosAneta ANNo ratings yet
- Wyklad 6 USG-1Document33 pagesWyklad 6 USG-1Amelia MotykaNo ratings yet
- Prezentacja PitagorasDocument10 pagesPrezentacja PitagorasReginaNo ratings yet
- Budowa Atomu7aDocument7 pagesBudowa Atomu7aIza ZiębaNo ratings yet
- Tworzenie Aplikacji Internetowych Z Użyciem Platformy Java EEDocument49 pagesTworzenie Aplikacji Internetowych Z Użyciem Platformy Java EEDevCastZoneNo ratings yet
- Kwasy Karboksylowe I PochodneDocument59 pagesKwasy Karboksylowe I Pochodneklaudynka6970No ratings yet