Download as pdf or txt
You might also like
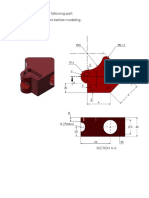
- Solidworks Cswa Exam Question #2Document14 pagesSolidworks Cswa Exam Question #2aldo0% (1)
- Joaquim A. Batlle, Ana Barjau Condomines - Rigid Body Kinematics-Cambridge University Press (2020)Document305 pagesJoaquim A. Batlle, Ana Barjau Condomines - Rigid Body Kinematics-Cambridge University Press (2020)vuliencnNo ratings yet
- StoneLight30, Brochure, 2016Document4 pagesStoneLight30, Brochure, 2016Servicio TécnicoNo ratings yet
- Major Revisions in Pancrustacean Phylogeny - 2023Document22 pagesMajor Revisions in Pancrustacean Phylogeny - 2023María JordánNo ratings yet
- Bronzati Et Al., 2023 Braincase and Neuroanatomy of The Lagerpetid Dromomeron GregoriiDocument28 pagesBronzati Et Al., 2023 Braincase and Neuroanatomy of The Lagerpetid Dromomeron Gregoriimauricio.garciaNo ratings yet
- Lynch.1982. Two New Species of Poison Dart Frogs - Edwardsi&ruiziDocument10 pagesLynch.1982. Two New Species of Poison Dart Frogs - Edwardsi&ruiziRaiza Nathaly Castañeda BonillaNo ratings yet
- Godoy 2014 Aplestosuchus Sordidus PDFDocument12 pagesGodoy 2014 Aplestosuchus Sordidus PDFFelipe MesVasNo ratings yet
- The Anatomical Record - 2024 - Desojo - Cranial Osteology and Paleoneurology of Tarjadia Ruthae An ErpetosuchidDocument35 pagesThe Anatomical Record - 2024 - Desojo - Cranial Osteology and Paleoneurology of Tarjadia Ruthae An ErpetosuchidCrocodilo VasconcelosNo ratings yet
- Ramirez & Grismado - RevMACN - 26 (1) 47-68 - 2024 (2) - Comprimido PDFDocument22 pagesRamirez & Grismado - RevMACN - 26 (1) 47-68 - 2024 (2) - Comprimido PDFSOSA CARDOZO JulietaNo ratings yet
- Unusual Looking Pentatomids: Reassessing The Taxonomy of Braunus Distant and Lojus McDonald (Hemiptera: Heteroptera: Pentatomidae)Document19 pagesUnusual Looking Pentatomids: Reassessing The Taxonomy of Braunus Distant and Lojus McDonald (Hemiptera: Heteroptera: Pentatomidae)bolosphex1634No ratings yet
- Structure, Innervation and Response Properties of Integumentary Sensory Organs in CrocodiliansDocument14 pagesStructure, Innervation and Response Properties of Integumentary Sensory Organs in CrocodiliansEduardo LeónNo ratings yet
- What Do Their Footprints Tell Us? Many Questions and Some Answers About The Life of Non Avian DinosaursDocument22 pagesWhat Do Their Footprints Tell Us? Many Questions and Some Answers About The Life of Non Avian DinosaursCrocodilo VasconcelosNo ratings yet
- Bolaños Dias Solé Zooming in On Amphibians Whis Is The Smallest Vertebrate in The World ZoologicaScripta 2024Document5 pagesBolaños Dias Solé Zooming in On Amphibians Whis Is The Smallest Vertebrate in The World ZoologicaScripta 2024bio.stephanoNo ratings yet
- Palaeontology - 2023 - Zhao - An Early Cambrian Polyp Reveals A Potential Anemone Like Ancestor For Medusozoan CnidariansDocument16 pagesPalaeontology - 2023 - Zhao - An Early Cambrian Polyp Reveals A Potential Anemone Like Ancestor For Medusozoan CnidarianskanishamqadaruNo ratings yet
- (2022) Müller Et Al. (Stenoscelida Aurantiacus)Document20 pages(2022) Müller Et Al. (Stenoscelida Aurantiacus)mauricio.garciaNo ratings yet
- Baiano - 2023 - Osteology of The Axial Skeleton of Aucasaurus Garridoi - Phylogenetic and Paleobiological InferencesDocument72 pagesBaiano - 2023 - Osteology of The Axial Skeleton of Aucasaurus Garridoi - Phylogenetic and Paleobiological InferencesplecotusNo ratings yet
- IS21076 CompressedDocument25 pagesIS21076 CompressedIkhva RiskianiNo ratings yet
- High Browsing Skeletal Adaptations in Spinophorosaurus Reveal An Evolutionary Innovation in Sauropod DinosaursDocument10 pagesHigh Browsing Skeletal Adaptations in Spinophorosaurus Reveal An Evolutionary Innovation in Sauropod DinosaursEdisonNo ratings yet
- 1242 Irr It at or OsteologyDocument117 pages1242 Irr It at or OsteologyJust JakeNo ratings yet
- Craniomandibular Anatomy of A Juvenile Specimen of Edmontosaurus Regalis Lambe 1917 Clarifies Issues in Ontogeny and BiogeographyDocument22 pagesCraniomandibular Anatomy of A Juvenile Specimen of Edmontosaurus Regalis Lambe 1917 Clarifies Issues in Ontogeny and BiogeographyCrocodilo VasconcelosNo ratings yet
- Bonvicino 2002Document11 pagesBonvicino 2002Maria EduardaNo ratings yet
- Journal of Anatomy - 2019 - Echeverr ADocument13 pagesJournal of Anatomy - 2019 - Echeverr Apjames199705No ratings yet
- VZ Article 76256 en 1Document18 pagesVZ Article 76256 en 1Erika CortesNo ratings yet
- Palopolo Et Al 2021 ZoroDocument18 pagesPalopolo Et Al 2021 ZoroGeorge ThoreauNo ratings yet
- A New Rhynchosaur (Reptilia, Archosauromorpha) From The Upper Triassic of Eastern North AmericaDocument12 pagesA New Rhynchosaur (Reptilia, Archosauromorpha) From The Upper Triassic of Eastern North AmericaPatriciaNo ratings yet
- Enigmatic Amphibians in Mid-Cretaceous Amber Were Chameleon-Like Ballistic FeedersDocument6 pagesEnigmatic Amphibians in Mid-Cretaceous Amber Were Chameleon-Like Ballistic FeedersDirtyBut WholeNo ratings yet
- The Anatomical Record - 2009 - Holliday - New Insights Into Dinosaur Jaw Muscle AnatomyDocument20 pagesThe Anatomical Record - 2009 - Holliday - New Insights Into Dinosaur Jaw Muscle AnatomyKidus TarekegnNo ratings yet
- Gonzalez Riga Et Al 2019Document43 pagesGonzalez Riga Et Al 2019Bernardo Javier Gonzalez RigaNo ratings yet
- App 20100005Document22 pagesApp 20100005Ghaier KazmiNo ratings yet
- Evolutionary Convergence in A Small Cursorial Styracosternan Ornithopod Dinosaur From Western Europe Calvarius RapidusDocument18 pagesEvolutionary Convergence in A Small Cursorial Styracosternan Ornithopod Dinosaur From Western Europe Calvarius RapidusMIGUEL ANGEL AVILA GUERRANo ratings yet
- A Four-Legged Snake From The Early Cretaceous of GondwanaDocument6 pagesA Four-Legged Snake From The Early Cretaceous of GondwanaYilbert Oswaldo Jimenez CanoNo ratings yet
- The Origin of The Turtle Body Plan EvideDocument19 pagesThe Origin of The Turtle Body Plan EvideXweuis Hekuos KweNo ratings yet
- Joa 13363Document15 pagesJoa 13363EdisonNo ratings yet
- Funtional Morphology of Bite Mechanics in The Great BarracudaDocument14 pagesFuntional Morphology of Bite Mechanics in The Great BarracudaassyahwaNo ratings yet
- Lambert 2016Document71 pagesLambert 2016Iqbal FitriantoNo ratings yet
- Vasquez-Restrepo and Diago-Toro 2023Document12 pagesVasquez-Restrepo and Diago-Toro 2023Diego BaldoNo ratings yet
- Molluscan Studies: Journal ofDocument19 pagesMolluscan Studies: Journal ofWidi SetyogatiNo ratings yet
- Ryan TaphonomyCentrosaurusOrnithischia 2001Document26 pagesRyan TaphonomyCentrosaurusOrnithischia 2001Crocodilo VasconcelosNo ratings yet
- Etymology: DimetrodonDocument5 pagesEtymology: DimetrodonAjay SinghNo ratings yet
- Biomecnica CocodriloDocument12 pagesBiomecnica CocodriloAngie Dayana Rivera BenavidesNo ratings yet
- Meraxes GigasDocument31 pagesMeraxes GigasGeronimo SampaolesiNo ratings yet
- Senaand Cubo 2023Document9 pagesSenaand Cubo 2023Crocodilo VasconcelosNo ratings yet
- Phylogenetic Inferences On The Systematics of Squaliform Sharks Based On Elasmobranch Scapular Morphology (Chondrichthyes: Elasmobranchii)Document18 pagesPhylogenetic Inferences On The Systematics of Squaliform Sharks Based On Elasmobranch Scapular Morphology (Chondrichthyes: Elasmobranchii)Karina SjNo ratings yet
- Strong Et Al 2020 - Insights Into Skull Evolution in Fossorial Snakes, As Revealed by The Cranial Morphology of Atractaspis IrregularisDocument27 pagesStrong Et Al 2020 - Insights Into Skull Evolution in Fossorial Snakes, As Revealed by The Cranial Morphology of Atractaspis IrregularisLivia PereiraNo ratings yet
- The Appendicular Skeleton of Suuwassea eDocument22 pagesThe Appendicular Skeleton of Suuwassea enomadNo ratings yet
- Macroevolutionary Patterns in The Pelvis StylopodiDocument48 pagesMacroevolutionary Patterns in The Pelvis StylopodiJorge MesoNo ratings yet
- Mayr Et Al-2016-PalaeontologyDocument11 pagesMayr Et Al-2016-PalaeontologyJorge BarNo ratings yet
- American Journal of Biological Anthropology - 2023 - Raventós Izard - The Morphofunctional Implications of The GlenoidDocument11 pagesAmerican Journal of Biological Anthropology - 2023 - Raventós Izard - The Morphofunctional Implications of The GlenoidIvana SovićNo ratings yet
- A Non-Avian Dinosaur With A Streamlined Body ExhibDocument9 pagesA Non-Avian Dinosaur With A Streamlined Body Exhibbraydenstoch777No ratings yet
- Texto CompletoDocument54 pagesTexto CompletoManuel PérezNo ratings yet
- Lateral Line Morphology and Cranial Osteology of The Rubynose Brotula, Cataetyx RubrirostrisDocument10 pagesLateral Line Morphology and Cranial Osteology of The Rubynose Brotula, Cataetyx RubrirostrisEloy Guillermo Mijares EchezuríaNo ratings yet
- Reconstrucción Zbrush Interesting Steyer Et Al 2010 3d ReconstructionsDocument9 pagesReconstrucción Zbrush Interesting Steyer Et Al 2010 3d ReconstructionsKisle VitaNo ratings yet
- A Sauropod From The Lower Jurassic La Quinta Formation (Dept. Cesar, Colombia) and The Initial Diversification of Eusauropods at Low LatitudesDocument25 pagesA Sauropod From The Lower Jurassic La Quinta Formation (Dept. Cesar, Colombia) and The Initial Diversification of Eusauropods at Low LatitudesUvuvwevwevwe Onyetenyevwe Ugwemuhwem OssasNo ratings yet
- Isasmendi Et Al 2024 - A New Basal Spinosaurid From The Enciso Group - OptimDocument2 pagesIsasmendi Et Al 2024 - A New Basal Spinosaurid From The Enciso Group - OptimJul JulNo ratings yet
- General Description: Triceratops Megalosaurus IguanodonDocument31 pagesGeneral Description: Triceratops Megalosaurus IguanodonRicardo Daniel Aranda ApolinarioNo ratings yet
- Phyletic Relationships of Living Sharks and RaysDocument20 pagesPhyletic Relationships of Living Sharks and RaysZAk1No ratings yet
- Taxonomic Revision of The Dasypus Kappleri Complex, With Revalidations of (2016)Document27 pagesTaxonomic Revision of The Dasypus Kappleri Complex, With Revalidations of (2016)sherazocallejasNo ratings yet
- Aschliman2012 Evolució de RayasDocument15 pagesAschliman2012 Evolució de RayasALHONDRA HORTENCIA GOMEZ SANCHEZNo ratings yet
- Systema Porifera PDFDocument20 pagesSystema Porifera PDFpeccerini100% (1)
- Müller & Garcia, 2022b, Oldest Dinosauromorpha South AmericaDocument7 pagesMüller & Garcia, 2022b, Oldest Dinosauromorpha South Americamauricio.garciaNo ratings yet
- The Lower Jurassic Ornithischian Dinosaur Cranial Anatomy, Functional Morphology, Taxonomy, and RelationshipsDocument95 pagesThe Lower Jurassic Ornithischian Dinosaur Cranial Anatomy, Functional Morphology, Taxonomy, and Relationshipsbraydenstoch777No ratings yet
- Chordate Evolution and The Three-Phylum System: ReviewDocument10 pagesChordate Evolution and The Three-Phylum System: Reviewnatalia_zimicz1412No ratings yet
- A New Genus of Pennsylvania Fish (Crossoperygii, Coelacanthiformes) from KansasFrom EverandA New Genus of Pennsylvania Fish (Crossoperygii, Coelacanthiformes) from KansasNo ratings yet
- Marsola Et Al. 2019 A New Dinosaur With Theropod Affinities From The Late Triassic Santa Maria Formation, South BrazilDocument25 pagesMarsola Et Al. 2019 A New Dinosaur With Theropod Affinities From The Late Triassic Santa Maria Formation, South Brazilmauricio.garciaNo ratings yet
- Pol Et Al. (2021) Triassic Sauropodomorph From South AmericaDocument14 pagesPol Et Al. (2021) Triassic Sauropodomorph From South Americamauricio.garciaNo ratings yet
- First Record of Stereospondyls Tetrapoda TemnosponDocument7 pagesFirst Record of Stereospondyls Tetrapoda Temnosponmauricio.garciaNo ratings yet
- Dias Da SilvaIlha2009Document7 pagesDias Da SilvaIlha2009mauricio.garciaNo ratings yet
- A New Typothoracine Aetosaur Archosauria Pseudosuchia From The Upper Triassic of India With Insights On Biostratigraphy Diversification and PaleoDocument23 pagesA New Typothoracine Aetosaur Archosauria Pseudosuchia From The Upper Triassic of India With Insights On Biostratigraphy Diversification and Paleomauricio.garciaNo ratings yet
- (2022) Müller & GarciaDocument21 pages(2022) Müller & Garciamauricio.garciaNo ratings yet
- Damke Et Al 2022 New LoricataDocument7 pagesDamke Et Al 2022 New Loricatamauricio.garciaNo ratings yet
- (2020) Müller & GarciaDocument5 pages(2020) Müller & Garciamauricio.garciaNo ratings yet
- Exam-Style Questions: E. B G B G × eDocument1 pageExam-Style Questions: E. B G B G × ePhan Minh ViệtNo ratings yet
- Combi Series 2023Document8 pagesCombi Series 2023KGC SaintifikNo ratings yet
- Motion in A Straight LineDocument16 pagesMotion in A Straight Lineuniverse2010No ratings yet
- H 1"" 1.9 Unfolding and Folding A ChainDocument4 pagesH 1"" 1.9 Unfolding and Folding A ChainAshNo ratings yet
- Lab Report 5 - Group 7 - Eh2203cDocument21 pagesLab Report 5 - Group 7 - Eh2203cAriff HaiqalNo ratings yet
- Dami SomoyeDocument26 pagesDami SomoyedamiNo ratings yet
- Adstefan Input Data SheetDocument4 pagesAdstefan Input Data SheetMuthu KumarNo ratings yet
- Product Design Based On Discrete Particle Modeling of A Uidized Bed GranulatorDocument12 pagesProduct Design Based On Discrete Particle Modeling of A Uidized Bed Granulator권혁록No ratings yet
- Bardon Initiation Into Hermetics (Better Outline) : Step X Exercises SummaryDocument7 pagesBardon Initiation Into Hermetics (Better Outline) : Step X Exercises SummaryRhaxma Conspiracy100% (1)
- Mock Test 1Document7 pagesMock Test 1abczyw.2006No ratings yet
- Principle of FlightDocument52 pagesPrinciple of FlightDang KhaiNo ratings yet
- DensityDocument24 pagesDensitytuvvacNo ratings yet
- International A Level Mathematics Pure Mathematics 3 Student Book SampleDocument34 pagesInternational A Level Mathematics Pure Mathematics 3 Student Book SampleHadi HindiNo ratings yet
- Singh 2010Document9 pagesSingh 2010annuNo ratings yet
- Mathematical ProofDocument4 pagesMathematical ProofsheridaNo ratings yet
- Materials: A Study On Tannery Sludge As A Raw Material For Cement MortarDocument14 pagesMaterials: A Study On Tannery Sludge As A Raw Material For Cement MortarKumar PallavNo ratings yet
- CH1 An Introduction To The Finite Element Method - v2Document21 pagesCH1 An Introduction To The Finite Element Method - v2Eng.Hesham AL-HelaleeNo ratings yet
- Science7 Q3 SLM12 PDFDocument14 pagesScience7 Q3 SLM12 PDFAlfred SolajesNo ratings yet
- Hydraulic Report Mudlogging ExampleDocument1 pageHydraulic Report Mudlogging ExampleCarmen Ibeth Olivos PradaNo ratings yet
- Week 7 q4Document7 pagesWeek 7 q4elizaldeNo ratings yet
- AphiweDocument16 pagesAphiweaphiweshozi18No ratings yet
- Agfa Optima 500Document20 pagesAgfa Optima 500j batlleNo ratings yet
- Door Frame (Choukat)Document21 pagesDoor Frame (Choukat)sirfmeinNo ratings yet
- Park Santamarina Revised Soil Classification SystemDocument14 pagesPark Santamarina Revised Soil Classification SystemPaul Alex Quiroz BarrionuevoNo ratings yet
- Physics Investigatory-Project-On-LdrDocument17 pagesPhysics Investigatory-Project-On-LdrSujay TewaryNo ratings yet
- Chrono PhysicsDocument2 pagesChrono PhysicsGaëtan de La VilleNo ratings yet
- Reservoir Geomechanics Sakessar ChorgaliDocument10 pagesReservoir Geomechanics Sakessar ChorgaliGhulam Mohyuddin SohailNo ratings yet