Download as pdf or txt
You might also like
- Budding Reader Book Set 1: Cat and RatFrom EverandBudding Reader Book Set 1: Cat and RatRating: 4.5 out of 5 stars4.5/5 (41)
- SITHCCC018 Prepare Food To Meet Special Dietary Requirements - Written Test V2.0Document26 pagesSITHCCC018 Prepare Food To Meet Special Dietary Requirements - Written Test V2.0Priyanka Thakur80% (5)
- Sri Lanka Tourism Case StudyDocument15 pagesSri Lanka Tourism Case StudyKirubaliny Prashath100% (3)
- Nutrition Assessment Subject McqsDocument13 pagesNutrition Assessment Subject McqsNoorNo ratings yet
- Brigade Calista BroacherDocument28 pagesBrigade Calista BroachereuphoriapropcornerNo ratings yet
- Orgl 3321 Dddmi ArtifactDocument5 pagesOrgl 3321 Dddmi Artifactapi-673975467No ratings yet
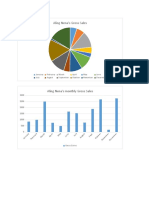
- Revenue by Month: BR Ua Ry M Ar CH Ap Ril M Ay Ju Ne Ju Ly Au Gu ST Se PT em Be R Oc To Be R No Ve M Be R de Ce M Be RDocument1 pageRevenue by Month: BR Ua Ry M Ar CH Ap Ril M Ay Ju Ne Ju Ly Au Gu ST Se PT em Be R Oc To Be R No Ve M Be R de Ce M Be Rkaneki kenNo ratings yet
- The Problem and A Review of Related LiteratureDocument36 pagesThe Problem and A Review of Related Literaturechariot1729No ratings yet
- Part 2 Fundamentals of Accountancy 1Document25 pagesPart 2 Fundamentals of Accountancy 1mang tomasNo ratings yet
- The Branches of Government: Lesson 1.2ADocument19 pagesThe Branches of Government: Lesson 1.2Apetermueller727No ratings yet
- Meaning Wheel With Citation 6Document1 pageMeaning Wheel With Citation 6Abu HanifahNo ratings yet
- Chart Title: Complaint Type No: of Complaints % Cumulative %Document2 pagesChart Title: Complaint Type No: of Complaints % Cumulative %Blossom KaurNo ratings yet
- Coke WaveDocument3 pagesCoke WavenikepeterNo ratings yet
- Presentation For August 15, 2012Document2 pagesPresentation For August 15, 2012elton jay amilaNo ratings yet
- Inspection Panel Compliance Advisor/Ombudsman: Unicate Corr Uption AilureDocument1 pageInspection Panel Compliance Advisor/Ombudsman: Unicate Corr Uption Ailureddlu123No ratings yet
- E 03 H 3 JobsDocument4 pagesE 03 H 3 JobsSarah Azizah WiratamaNo ratings yet
- Austerlitz Boulogne Pont de Saint - CloudDocument1 pageAusterlitz Boulogne Pont de Saint - CloudtudoseNo ratings yet
- Criteria For Judging ClassroomDocument2 pagesCriteria For Judging ClassroomMaritessNo ratings yet
- Crawford County Drug Overdose Fatality Review's 2021 ReportDocument13 pagesCrawford County Drug Overdose Fatality Review's 2021 ReportGere Goble0% (1)
- Mediterranean SeaDocument6 pagesMediterranean SeaJonahNo ratings yet
- Results From The DORA Survey2Document12 pagesResults From The DORA Survey2Myroslava BoichukNo ratings yet
- Ingredient Vulnerability Assessment BRCDocument3 pagesIngredient Vulnerability Assessment BRCSherylNo ratings yet
- Organelles PracticeDocument65 pagesOrganelles PracticeAlfaNo ratings yet
- ActivityForTheDay 03Document1 pageActivityForTheDay 03shamimabegam1981No ratings yet
- Kalender Pameran-2Document1 pageKalender Pameran-2Adwan Dwi SasonoNo ratings yet
- Group 9 - Oil and Gas IndustryDocument25 pagesGroup 9 - Oil and Gas IndustrymohilNo ratings yet
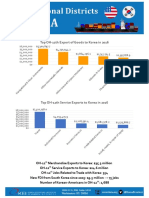
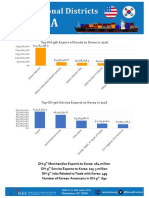
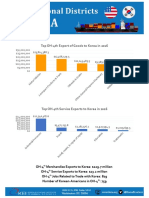
- Top OH-12th Export of Goods To Korea in 2016Document2 pagesTop OH-12th Export of Goods To Korea in 2016Korea Economic Institute of America (KEI)No ratings yet
- Top OH-9th Export of Goods To Korea in 2016Document2 pagesTop OH-9th Export of Goods To Korea in 2016Korea Economic Institute of America (KEI)No ratings yet
- Program Uks: Oleh Hj. Suhartini, S. KepDocument3 pagesProgram Uks: Oleh Hj. Suhartini, S. KepSuhartini KhumairahNo ratings yet
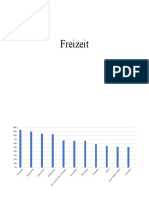
- FreizeitDocument2 pagesFreizeitZille Tóth-NémethNo ratings yet
- Marshmallow TowerDocument1 pageMarshmallow TowerTarcayani PrebagaranNo ratings yet
- Aggregate PlanningDocument85 pagesAggregate PlanninggmbhuvaneswariNo ratings yet
- Excel Tutorial Name Arnold Byun Teacher MR - Heil Class/Period Ca0 Date 7/3/2010Document6 pagesExcel Tutorial Name Arnold Byun Teacher MR - Heil Class/Period Ca0 Date 7/3/2010absangwonNo ratings yet
- Opol National Secondary Technical School Opol, Misamis Oriental Progress ChartDocument3 pagesOpol National Secondary Technical School Opol, Misamis Oriental Progress ChartChariss Joy LacayaNo ratings yet
- Knowledge Gateway PresentationDocument33 pagesKnowledge Gateway PresentationMunhzorig BattulgaNo ratings yet
- Exercise 1: Looking For Information at SpeedDocument17 pagesExercise 1: Looking For Information at SpeedtruongthikhaNo ratings yet
- Bad PDFCreatorDocument3 pagesBad PDFCreatorapi-3766300100% (2)
- Aling Nena's Gross Sales: January February March April May June July Augost September October November DecemberDocument1 pageAling Nena's Gross Sales: January February March April May June July Augost September October November DecemberJerry G. GabacNo ratings yet
- 55265A - Microsoft PowerApps - Skillpipe - Mod - 1Document19 pages55265A - Microsoft PowerApps - Skillpipe - Mod - 1Desarrollo Gran Colombia Gold100% (1)
- 55265A - Microsoft PowerApps - Skillpipe - Mod - 4Document15 pages55265A - Microsoft PowerApps - Skillpipe - Mod - 4Desarrollo Gran Colombia GoldNo ratings yet
- Job Situation in The Recession: April 2009Document34 pagesJob Situation in The Recession: April 2009Khawaja_FaisalNo ratings yet
- Session 1 SVDocument50 pagesSession 1 SVfelinethienpontNo ratings yet
- Chuck e Cheese NutritionalsDocument13 pagesChuck e Cheese NutritionalsdengnisiphotosNo ratings yet
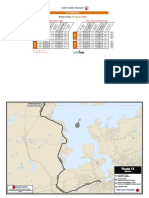
- Route 12 Martinon May32015Document2 pagesRoute 12 Martinon May32015Vy Thoai0% (1)
- Hypnobirthing Study AustraliaDocument7 pagesHypnobirthing Study AustraliaRia Ulfha IndrianyNo ratings yet
- Moloi Peter M: RN Am e Na M e Ge ND Er en GL Ish Zu Lu Hi ST or y SC Ie NC e Bi Ol Og y Av Er Ag e Re Su LTDocument4 pagesMoloi Peter M: RN Am e Na M e Ge ND Er en GL Ish Zu Lu Hi ST or y SC Ie NC e Bi Ol Og y Av Er Ag e Re Su LTtsangwani_mudzunga emilyNo ratings yet
- Top OH-4th Export of Goods To Korea in 2016Document2 pagesTop OH-4th Export of Goods To Korea in 2016Korea Economic Institute of America (KEI)No ratings yet
- Top OH-7th Export of Goods To Korea in 2016Document2 pagesTop OH-7th Export of Goods To Korea in 2016Korea Economic Institute of America (KEI)No ratings yet
- 10 SCL TaposDocument44 pages10 SCL TaposRoss Elmar OrolfoNo ratings yet
- Section5 1-HW1Document5 pagesSection5 1-HW1Jeremy SydorNo ratings yet
- 55265A - Microsoft PowerApps - Skillpipe - Mod - 2Document22 pages55265A - Microsoft PowerApps - Skillpipe - Mod - 2Desarrollo Gran Colombia Gold100% (1)
- Food Delivery DFDDocument8 pagesFood Delivery DFDofficialigamerytNo ratings yet
- Class 5 114221022Document3 pagesClass 5 114221022Md. Abdulla Al AhadNo ratings yet
- IBR Editorial CalendarDocument12 pagesIBR Editorial CalendarEllerique ShamsuddinNo ratings yet
- Describing Bar Chart Practice 3Document7 pagesDescribing Bar Chart Practice 3Irfan HasifNo ratings yet
- Índice: Jorge Augusto Llamas Caamaño (jorgeฺllamas@opxisฺcom) has a non-transferable license to use this Student GuideฺDocument12 pagesÍndice: Jorge Augusto Llamas Caamaño (jorgeฺllamas@opxisฺcom) has a non-transferable license to use this Student GuideฺRonald LLamasNo ratings yet
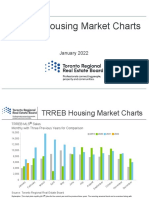
- Market-Stats Housing-Charts TREB Housing Market Charts-January 2022Document9 pagesMarket-Stats Housing-Charts TREB Housing Market Charts-January 2022gul khaNo ratings yet
- 02-Sciencethe Unexplored BiosphereDocument21 pages02-Sciencethe Unexplored BiospherebiostatisticsNo ratings yet
- Tms Risk Assessment EmergencyDocument1 pageTms Risk Assessment Emergencyapi-703370744No ratings yet
- Smart Box 16x9Document52 pagesSmart Box 16x9German RojasNo ratings yet
- To EDIT GRAPH CSPMAP - PIR Presentation (Rizal Medical Center) 2023Document29 pagesTo EDIT GRAPH CSPMAP - PIR Presentation (Rizal Medical Center) 2023Oncology PharmacyNo ratings yet
- Afectiuni Produse Mod de Administrare: Indicatii Terapeutice Cu Produsele Complementare Forever Living ProductsDocument13 pagesAfectiuni Produse Mod de Administrare: Indicatii Terapeutice Cu Produsele Complementare Forever Living Productsluminitalasc100% (1)
- De Thi Anh CHUYEN TPHCM 2018 Va DapAnDocument6 pagesDe Thi Anh CHUYEN TPHCM 2018 Va DapAnTruong TranNo ratings yet
- MALUNGGAY (Moringa Oleifera) EthnobotanicalDocument4 pagesMALUNGGAY (Moringa Oleifera) EthnobotanicalDez TabiosNo ratings yet
- Applied Nutrition: Prof. Mona HegazyDocument150 pagesApplied Nutrition: Prof. Mona HegazyChristina AtefNo ratings yet
- Eating For Your Blood Type - AB+ & AB - Diet TipsDocument6 pagesEating For Your Blood Type - AB+ & AB - Diet TipsJonathan QuilacioNo ratings yet
- Bio XI NutritionDocument2 pagesBio XI NutritionAbdul Haseeb JokhioNo ratings yet
- 10 I January 2022Document7 pages10 I January 2022IJRASETPublicationsNo ratings yet
- Harga HerbalifeDocument4 pagesHarga Herbalifesunarni100% (1)
- BIOC201 Tutorial 2 (2024)Document1 pageBIOC201 Tutorial 2 (2024)hlongwanasnenhlanhla8No ratings yet
- Albizia Saman 3Document3 pagesAlbizia Saman 3Jesus Llorente MendozaNo ratings yet
- Diets Modified in CompositionDocument6 pagesDiets Modified in Compositionsy saludNo ratings yet
- 0610 - 43 BIOLOGY November 2019 Theory ExtendedDocument8 pages0610 - 43 BIOLOGY November 2019 Theory ExtendedManali Mehta ParikhNo ratings yet
- Clinical 1 Case Study 1Document6 pagesClinical 1 Case Study 1api-535299330No ratings yet
- 30-Second NutritionDocument163 pages30-Second NutritionRahul Roy100% (1)
- 10 Tips Healthy LifestyleDocument2 pages10 Tips Healthy LifestyleGeorge AbuladzeNo ratings yet
- Examen Unidades 6 y 8Document5 pagesExamen Unidades 6 y 8joseNo ratings yet
- Faq Protein PDFDocument3 pagesFaq Protein PDFNikita ZhukNo ratings yet
- Activity 9 BiochemDocument5 pagesActivity 9 BiochemElvie BejecNo ratings yet
- Nutdth Module Rda ReniDocument7 pagesNutdth Module Rda ReniKwebblekop JordiNo ratings yet
- Casein in Different Samples of MilkDocument23 pagesCasein in Different Samples of MilkRAJALAKSHMI SNo ratings yet
- Easy and Quick Chicken Cat Food RecipeDocument8 pagesEasy and Quick Chicken Cat Food RecipeinsiderrebornNo ratings yet
- Lecture (9) Integration of MetabolismDocument40 pagesLecture (9) Integration of MetabolismMahmoud ElazbNo ratings yet
- Term-1 MCQ 3Document5 pagesTerm-1 MCQ 3Ansh KumarNo ratings yet
- Wild Edible Plants As A Food Resource: Traditional KnowledgeDocument20 pagesWild Edible Plants As A Food Resource: Traditional KnowledgeMikeNo ratings yet
- L01 Nutrifinds Wholesale PricelistDocument10 pagesL01 Nutrifinds Wholesale PricelistPaul LechugaNo ratings yet
- DPP - 01 Biomolecule Carbohydrate & ProteinsDocument2 pagesDPP - 01 Biomolecule Carbohydrate & ProteinsBIONEETI BIONEETINo ratings yet
- Statement of Account: DMM'S Catering ServicesDocument2 pagesStatement of Account: DMM'S Catering ServicesJunreyboi RemixNo ratings yet
- ATM - GA - 2 - Pivot Table - SOLUTIONDocument52 pagesATM - GA - 2 - Pivot Table - SOLUTIONSuraj Kumar BarnwalNo ratings yet