Download as pdf or txt
You might also like
- Business and Society Stakeholders Ethics Public Policy 17Th Edition Anne Lawrence Full ChapterDocument67 pagesBusiness and Society Stakeholders Ethics Public Policy 17Th Edition Anne Lawrence Full Chaptermichael.lynch15578% (9)
- Hi-Survey Road User Manual-EN-20200330Document276 pagesHi-Survey Road User Manual-EN-20200330Mauricio RuizNo ratings yet
- Checklist Audit ISO 22000Document18 pagesChecklist Audit ISO 22000Abhishek Kumar Singh95% (20)
- ESPECIFICACIONES D12C Motor VolvoDocument44 pagesESPECIFICACIONES D12C Motor VolvoErika Salinas100% (4)
- Deconstructing Cortical Folding - Genetic, Cellular and Mechanical DeterminantsDocument16 pagesDeconstructing Cortical Folding - Genetic, Cellular and Mechanical DeterminantsAndriel CardosoNo ratings yet
- Trim Clean 2030-Msds (Bld-1656 v1)Document8 pagesTrim Clean 2030-Msds (Bld-1656 v1)Tri Febrianto PamungkasNo ratings yet
- Mitosis Through Microscope Science 2003Document7 pagesMitosis Through Microscope Science 2003Fran IgarzabalNo ratings yet
- CAREW, T. J. - Startle Circuits - Neural Mechanisms of Startle Behavior. (1985) (10.1126 - Science.229.4709.158) - Libgen - LiDocument2 pagesCAREW, T. J. - Startle Circuits - Neural Mechanisms of Startle Behavior. (1985) (10.1126 - Science.229.4709.158) - Libgen - LinajiNo ratings yet
- Genetics, Cell Cycle and Cell Expansion in Organogenesis in PlantsDocument4 pagesGenetics, Cell Cycle and Cell Expansion in Organogenesis in PlantsNaincy ChandanNo ratings yet
- 3769 FullDocument12 pages3769 FullAgnes GintingNo ratings yet
- Nihms 367187Document14 pagesNihms 367187Tiego FerreiraNo ratings yet
- Three-Dimensional Visualization of The Human Membranous Labyrinth - The Membrana Limitans and Its Role in Vestibular FormDocument15 pagesThree-Dimensional Visualization of The Human Membranous Labyrinth - The Membrana Limitans and Its Role in Vestibular FormManuek GarciaNo ratings yet
- Craig and Banker 1994Document44 pagesCraig and Banker 1994Kanwal RashidNo ratings yet
- Did Consciousness Cause The Cambrian Evolutionary ExplosionDocument18 pagesDid Consciousness Cause The Cambrian Evolutionary ExplosionBerry BernieNo ratings yet
- Museum of Comparative Zoology, Harvard University, Cambridge, Massachusetts 02138Document12 pagesMuseum of Comparative Zoology, Harvard University, Cambridge, Massachusetts 02138Julio César Chávez GalarzaNo ratings yet
- The Minicolumn Hypothesis in NeuroscienceDocument18 pagesThe Minicolumn Hypothesis in NeuroscienceRodrigo SanzNo ratings yet
- 01 CiliaDocument3 pages01 CiliaLaksh NrusimhanNo ratings yet
- Hochner2006 - Octopus BrainDocument11 pagesHochner2006 - Octopus BrainGuilherme SouzaNo ratings yet
- A Long Twentieth Century of The Cell Cycle and BeyDocument8 pagesA Long Twentieth Century of The Cell Cycle and BeyVidhi SaxenaNo ratings yet
- Dynamics of Intestinal PropulsionDocument17 pagesDynamics of Intestinal PropulsionLDaggersonNo ratings yet
- Development and Plasticity of The Corpus CallosumDocument15 pagesDevelopment and Plasticity of The Corpus CallosumOmelas OmNo ratings yet
- The Actin Cortex at A GlanceDocument9 pagesThe Actin Cortex at A GlanceArkayan LahaNo ratings yet
- Taylor 1974Document31 pagesTaylor 1974Renato AmosNo ratings yet
- The Minicolumn Hypothesis in NeuroscienceDocument17 pagesThe Minicolumn Hypothesis in NeuroscienceAlyssa LeonidoNo ratings yet
- Art 01Document12 pagesArt 01nicolasNo ratings yet
- Ler Da Thais Reprodução 2Document11 pagesLer Da Thais Reprodução 2Gustavo FelpeNo ratings yet
- Vertebrates: Comparative Anatomy, Function, Evolution. - Kenneth V. Kardong. 1998. Second Edition. Mcgraw-Hill, Boston, MassachusettsDocument3 pagesVertebrates: Comparative Anatomy, Function, Evolution. - Kenneth V. Kardong. 1998. Second Edition. Mcgraw-Hill, Boston, MassachusettsMinnie chanNo ratings yet
- Evolutionary Consequences of Skeletal DifferentiationDocument22 pagesEvolutionary Consequences of Skeletal Differentiationjulio7345No ratings yet
- PIIS0092867405002333Document3 pagesPIIS0092867405002333Who ?No ratings yet
- Introduction To Insect Sensory Organs As A Model System in Sensory Physiology and Developmental BiologyDocument3 pagesIntroduction To Insect Sensory Organs As A Model System in Sensory Physiology and Developmental BiologypajtgocjNo ratings yet
- Cold Spring Harb Perspect Biol-2014-BurkiDocument19 pagesCold Spring Harb Perspect Biol-2014-BurkiCarmen CaballeroNo ratings yet
- TMP EAB5Document6 pagesTMP EAB5FrontiersNo ratings yet
- The Differential Roles of Periosteal and Capsular Functional Matrices in Orofacial GrowthDocument6 pagesThe Differential Roles of Periosteal and Capsular Functional Matrices in Orofacial GrowthCony Del CidNo ratings yet
- Wiley Taxon: This Content Downloaded From 197.255.118.131 On Thu, 04 Apr 2019 12:25:04 UTCDocument31 pagesWiley Taxon: This Content Downloaded From 197.255.118.131 On Thu, 04 Apr 2019 12:25:04 UTCSaheed AbdulkarimNo ratings yet
- The Forces Behind Cell Movement: Revathi Ananthakrishnan and Allen EhrlicherDocument15 pagesThe Forces Behind Cell Movement: Revathi Ananthakrishnan and Allen EhrlicherPicha PichiNo ratings yet
- Journal FreeDocument36 pagesJournal FreeRiccardo Spinoza SilvaNo ratings yet
- Origin of The Cell Nucleus, Mitosis and Sex Roles of Intracellular CoevolutionDocument78 pagesOrigin of The Cell Nucleus, Mitosis and Sex Roles of Intracellular CoevolutionnolosupeNo ratings yet
- Anderson, 2006 PDFDocument17 pagesAnderson, 2006 PDFMario GomezNo ratings yet
- Esposito 2014Document19 pagesEsposito 2014maria.pezzottiNo ratings yet
- Earth and Life 2Document5 pagesEarth and Life 2ashleydemiar7No ratings yet
- Cell Theory RevisedDocument24 pagesCell Theory RevisedCésar Octavio Moreno ZúñigaNo ratings yet
- (Book) Welch, Clegg, 1985 - The Organization of Cell Metabolism PDFDocument373 pages(Book) Welch, Clegg, 1985 - The Organization of Cell Metabolism PDFBogdan HateganNo ratings yet
- Motor Axon Regeneration and Muscle Reinnervation in Young Adult and Aged AnimalsDocument12 pagesMotor Axon Regeneration and Muscle Reinnervation in Young Adult and Aged AnimalsIsaac AbadNo ratings yet
- Szabó, Mayor - 2015 - Cell Traction in Collective Cell Migration and Morphogenesis The Chase and Run MechanismDocument5 pagesSzabó, Mayor - 2015 - Cell Traction in Collective Cell Migration and Morphogenesis The Chase and Run Mechanismlpa.ufmt.enfNo ratings yet
- Hashiguchi Et Al 2013 - Gravity Sensing Article - ImagenesDocument10 pagesHashiguchi Et Al 2013 - Gravity Sensing Article - ImagenesArmando Irisarri AmatriainNo ratings yet
- Trager 1963Document6 pagesTrager 1963IñigoGaitanSalvatellaNo ratings yet
- SynaptogenesisDocument5 pagesSynaptogenesisRitesh BawriNo ratings yet
- Eukaryotic Cells and Their Cell Bodies (Baluska Et Al., 2004)Document24 pagesEukaryotic Cells and Their Cell Bodies (Baluska Et Al., 2004)ej valerioNo ratings yet
- Caveolin-Deficient Mice: Insights Into Caveolar Function Human DiseaseDocument10 pagesCaveolin-Deficient Mice: Insights Into Caveolar Function Human DiseaseStefania CristinaNo ratings yet
- Agenesis of The Corpus Callosum: Genetic, Developmental and Functional Aspects of ConnectivityDocument14 pagesAgenesis of The Corpus Callosum: Genetic, Developmental and Functional Aspects of ConnectivityAyu PratiwiNo ratings yet
- Biology Checklist From TextbookDocument12 pagesBiology Checklist From Textbookrayaan.joshiNo ratings yet
- Naturalizing Values: Organisms and Species by Holmes Rolston IIIDocument14 pagesNaturalizing Values: Organisms and Species by Holmes Rolston IIICenter for Respect of Life and EnvironmentNo ratings yet
- ASSIGNMENT CC2 SPECIAL - Autosaved - PDFDocument70 pagesASSIGNMENT CC2 SPECIAL - Autosaved - PDFDEVDEEP SIKDARNo ratings yet
- Desarrollo Del CloroplastoDocument21 pagesDesarrollo Del CloroplastoAnderson HernándezNo ratings yet
- The Chicken As A Model For Embryonic Development: M.G. Davey C. TickleDocument9 pagesThe Chicken As A Model For Embryonic Development: M.G. Davey C. TickleUtari Septiana DewiNo ratings yet
- Olfaction, Navigation, and The Origin of Isocortex: Francisco Aboitiz and Juan F. MontielDocument12 pagesOlfaction, Navigation, and The Origin of Isocortex: Francisco Aboitiz and Juan F. MontielmioclaenusNo ratings yet
- Introductory Discussion On Glial Function: Department of Psychology, Yale University, New Haven, Conn. U.S.A.)Document11 pagesIntroductory Discussion On Glial Function: Department of Psychology, Yale University, New Haven, Conn. U.S.A.)Eusebio ChaconNo ratings yet
- Cortical Activity Early Brain DevelopDocument12 pagesCortical Activity Early Brain DevelopDa CuNo ratings yet
- Review Tubulin-Dynein System in Agellar and Ciliary MovementDocument19 pagesReview Tubulin-Dynein System in Agellar and Ciliary Movementsama EmadNo ratings yet
- The Differential Roles of Periosteal and Capsular Functional Matrices in Orofacial GrowthDocument6 pagesThe Differential Roles of Periosteal and Capsular Functional Matrices in Orofacial GrowthAaron Neal DmdNo ratings yet
- Developmental Biology: Juliana M. Coelho-Aguiar, Nicole M. Le Douarin, Elisabeth DupinDocument13 pagesDevelopmental Biology: Juliana M. Coelho-Aguiar, Nicole M. Le Douarin, Elisabeth DupinCarlos PalhaNo ratings yet
- DrosophiliaDocument6 pagesDrosophiliaenoch taclanNo ratings yet
- 25 VogelDocument12 pages25 VogelThati PonceNo ratings yet
- The Ocular Lens: A Classic Model For Development, Physiology and DiseaseDocument3 pagesThe Ocular Lens: A Classic Model For Development, Physiology and DiseaseWahyudi Permana DarlisNo ratings yet
- Evolution: What Darwin Did Not Know by Then..! [And the Origin of Species Through Species-Branding]From EverandEvolution: What Darwin Did Not Know by Then..! [And the Origin of Species Through Species-Branding]No ratings yet
- 561566642977unit-3 - TextilesDocument11 pages561566642977unit-3 - TextilesGaganpreet Kaur Fashion DesigningNo ratings yet
- GSB PDFDocument4 pagesGSB PDFsachin sNo ratings yet
- DNV-CG-0037 2021-11Document74 pagesDNV-CG-0037 2021-11wfxNo ratings yet
- Living in The I.T Era Module 2Document18 pagesLiving in The I.T Era Module 2Aivee GatzzNo ratings yet
- Cancer and Its Easy Treatment in Homeopathy - Bashir Mahmud ElliasDocument5 pagesCancer and Its Easy Treatment in Homeopathy - Bashir Mahmud ElliasBashir Mahmud ElliasNo ratings yet
- Maya Under Water LightingDocument12 pagesMaya Under Water LightingKombiah RkNo ratings yet
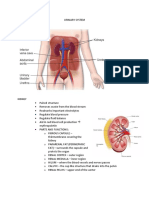
- Urinary SystemDocument9 pagesUrinary SystemMary Joyce RamosNo ratings yet
- BCGS MP-86Document511 pagesBCGS MP-86Valentina Renza PinoNo ratings yet
- Microcontroller Based Speech To Text Translation SystemDocument6 pagesMicrocontroller Based Speech To Text Translation SystemJay Prakash MandalNo ratings yet
- P2 Chp2 Section 2.1ADocument3 pagesP2 Chp2 Section 2.1APaing Khant KyawNo ratings yet
- Unit Two. Dominant Theories Approaches: Positivism Rational Choice Theory MarxismDocument8 pagesUnit Two. Dominant Theories Approaches: Positivism Rational Choice Theory MarxismPaul RicoNo ratings yet
- Test Report CMI, 200 Amp 35kV Class "Tuf-Ex-Well II" Bushing WellDocument8 pagesTest Report CMI, 200 Amp 35kV Class "Tuf-Ex-Well II" Bushing WellCristobal BohorquezNo ratings yet
- GCSE-Biology-7-Summer2019-Foundation Tier, Unit 1-PaperDocument24 pagesGCSE-Biology-7-Summer2019-Foundation Tier, Unit 1-PaperjramatlhakolaneNo ratings yet
- Lab ManualDocument41 pagesLab Manualprojectapply2023No ratings yet
- Especificaciones de Motor j15 de Datsun NissanDocument2 pagesEspecificaciones de Motor j15 de Datsun NissanPablo Andres60% (5)
- The Talmud of Jerusalem - Schwab Moise 1839-1918 TRDocument201 pagesThe Talmud of Jerusalem - Schwab Moise 1839-1918 TRJanice O'BrianNo ratings yet
- BS Soil and Rock Explanatory NotesDocument15 pagesBS Soil and Rock Explanatory NotesvictorNo ratings yet
- Feasibility Study of New Zealand Radiata Pine Cross-Laminated TimberDocument5 pagesFeasibility Study of New Zealand Radiata Pine Cross-Laminated TimberClaudio VillegasNo ratings yet
- ARTS7 Q3 Week6Document7 pagesARTS7 Q3 Week6Samantha Nicole CarlotoNo ratings yet
- Product Data: Order Tracking Analyzer - Type 2145Document8 pagesProduct Data: Order Tracking Analyzer - Type 2145jhon vargasNo ratings yet
- Controllers and Controller StationsDocument65 pagesControllers and Controller StationsLucian ChorusNo ratings yet
- Assignment2 NamocDocument5 pagesAssignment2 NamocHenry Darius NamocNo ratings yet
- Individual Development WorkoutDocument3 pagesIndividual Development WorkoutmichelleNo ratings yet
- PSC Unit 3Document18 pagesPSC Unit 3Santosh ReddyNo ratings yet
- Cryolite JM File 2011Document5 pagesCryolite JM File 2011mutemuNo ratings yet
































































![Evolution: What Darwin Did Not Know by Then..! [And the Origin of Species Through Species-Branding]](https://imgv2-1-f.scribdassets.com/img/word_document/270231362/149x198/aea4885cd2/1677109978?v=1)