
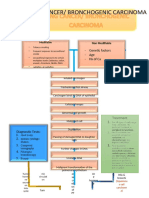
HLA-B27 Associated Spondyloarthropathy, An
HLA-B27 Associated Spondyloarthropathy, An
You might also like
- Elite 100 NBME Concepts OutlineDocument158 pagesElite 100 NBME Concepts OutlineDeborah Peters100% (3)
- Chapter 36: Aerosol Drug Therapy Test Bank Multiple ChoiceDocument25 pagesChapter 36: Aerosol Drug Therapy Test Bank Multiple ChoiceKhalid Julkanain50% (2)
- Morphine Mikhail BulgakovDocument23 pagesMorphine Mikhail Bulgakovciuciu.oxanaNo ratings yet
- HBsAg WantaiDocument16 pagesHBsAg WantaiAnnisa UtamiNo ratings yet
- Rheumatology Cases Fall Clinics 2019Document85 pagesRheumatology Cases Fall Clinics 2019NaziBrola TsivadzeNo ratings yet
- Ultimate Stretching GuideDocument18 pagesUltimate Stretching GuideJason Collingburn100% (11)
- Chiropractic Technique Self Adjustment Made Easy PDFDocument40 pagesChiropractic Technique Self Adjustment Made Easy PDFEashwar Prasad Meenakshi100% (3)
- Mcmichael 2002Document6 pagesMcmichael 2002peppa pigNo ratings yet
- Does HLA-B27 Status Influence AnkylosingDocument6 pagesDoes HLA-B27 Status Influence AnkylosingcismarcivNo ratings yet
- A Noncompeting Pair of Human Neutralizing Antibodies Block COVID-19 Virus Binding To Its Receptor ACE2Document6 pagesA Noncompeting Pair of Human Neutralizing Antibodies Block COVID-19 Virus Binding To Its Receptor ACE2Walid BoukhelkhalNo ratings yet
- Reactive Arthritis From Clinical Features To PathoDocument11 pagesReactive Arthritis From Clinical Features To Patho[161]Shuaib AktherNo ratings yet
- Bowness 2015 Hla b27Document22 pagesBowness 2015 Hla b27jezy2014No ratings yet
- HLA-B27 Is A Risk Factor For Rheumatoid Arthritis Suggestion ForDocument4 pagesHLA-B27 Is A Risk Factor For Rheumatoid Arthritis Suggestion ForcismarcivNo ratings yet
- COVID-19 Vaccine Design: The Janus Face of Immune EnhancementDocument2 pagesCOVID-19 Vaccine Design: The Janus Face of Immune EnhancementPratiwi PuspanegaraNo ratings yet
- HTTPS:WWW Ncbi NLM Nih gov:pmc:articles:PMC8040313:pdf:ijbsv17p1125Document13 pagesHTTPS:WWW Ncbi NLM Nih gov:pmc:articles:PMC8040313:pdf:ijbsv17p1125Ei DrakorNo ratings yet
- Covid Mda5 Jo1 Mi2Document15 pagesCovid Mda5 Jo1 Mi2Raul ReyesNo ratings yet
- 534 FullDocument9 pages534 FullSoehalino HalNo ratings yet
- Voruganti and Bowness 2020 - New Developments in Our Understanding of Ankylosing Spondylitis Patho-AnnotatedDocument9 pagesVoruganti and Bowness 2020 - New Developments in Our Understanding of Ankylosing Spondylitis Patho-AnnotatedmuamarrayNo ratings yet
- New Pathogenic Hypotheses For SpondyloarthropathiesDocument9 pagesNew Pathogenic Hypotheses For SpondyloarthropathiesAli ChaconNo ratings yet
- Parvovirus Lupus PDFDocument3 pagesParvovirus Lupus PDFDaniel PintoNo ratings yet
- Kel 329Document5 pagesKel 329dr.rahafturkiNo ratings yet
- How To Interpret and Use COVID-19 Serology and Immunology TestsDocument6 pagesHow To Interpret and Use COVID-19 Serology and Immunology TestsDana MNo ratings yet
- Science Abc2241 FullDocument8 pagesScience Abc2241 FullEconomy 365No ratings yet
- ABO Blood Group Type Does Not Influence The LevelDocument8 pagesABO Blood Group Type Does Not Influence The LevelANANo ratings yet
- Bowness 2015Document22 pagesBowness 2015shijumon8055No ratings yet
- Comment: The Potential Danger of Suboptimal Antibody Responses in COVID-19Document3 pagesComment: The Potential Danger of Suboptimal Antibody Responses in COVID-19Flavia Oliveira PampolhaNo ratings yet
- Genetics of Axial SpondyloarthritisDocument1 pageGenetics of Axial SpondyloarthritiscismarcivNo ratings yet
- HLA-B27 What's NewDocument11 pagesHLA-B27 What's NewcismarcivNo ratings yet
- Aps Etiologia PatogenesisDocument21 pagesAps Etiologia PatogenesisLeonardo GarciaNo ratings yet
- Ankylosing Spondylitis HLA-B27 and Klebsiella - AnDocument32 pagesAnkylosing Spondylitis HLA-B27 and Klebsiella - AnbciebienNo ratings yet
- HLA-C An Accomplice in Rheumatic DiseasesDocument9 pagesHLA-C An Accomplice in Rheumatic DiseasesNaziBrola TsivadzeNo ratings yet
- s12026 018 9045 0Document10 pagess12026 018 9045 0ndsjNo ratings yet
- Regular Article: Concurrent in Uenza Vaccination Reduces anti-FVIII Antibody Responses in Murine Hemophilia ADocument11 pagesRegular Article: Concurrent in Uenza Vaccination Reduces anti-FVIII Antibody Responses in Murine Hemophilia Ahasemana hasemanaNo ratings yet
- Review Article: Pathogenic and Epiphenomenal Anti-DNA Antibodies in SLEDocument18 pagesReview Article: Pathogenic and Epiphenomenal Anti-DNA Antibodies in SLEvishnupgiNo ratings yet
- Progress in Guillain Barr Syndrome Immunotherapy A Narrative Review of New Strategies in Recent YearsDocument9 pagesProgress in Guillain Barr Syndrome Immunotherapy A Narrative Review of New Strategies in Recent YearsReem YousifNo ratings yet
- PIIS2589537023003310Document10 pagesPIIS25895370230033109jz5dzj5gwNo ratings yet
- Richardson 2017Document8 pagesRichardson 2017YogiAnantaNo ratings yet
- Cei0121 0270Document5 pagesCei0121 0270Nur WahyuniNo ratings yet
- A Comprehensive Review of Biological Agents For Lupus Beyond Single TargetDocument11 pagesA Comprehensive Review of Biological Agents For Lupus Beyond Single TargetBelen LemaNo ratings yet
- The SARS-Coronavirus-Host Interactome: Identification of Cyclophilins As Target For Pan-Coronavirus InhibitorsDocument15 pagesThe SARS-Coronavirus-Host Interactome: Identification of Cyclophilins As Target For Pan-Coronavirus Inhibitorsfaris daboolNo ratings yet
- Vaccination Versus Hybrid ImmunizationDocument9 pagesVaccination Versus Hybrid ImmunizationJovanaNo ratings yet
- B Cell Responses To HIV InfectionDocument30 pagesB Cell Responses To HIV InfectionLhorruama DiasNo ratings yet
- The Etiological Role of Chlamydia Pneumoniae and Erythematosus of Iraqi Female PatientsDocument5 pagesThe Etiological Role of Chlamydia Pneumoniae and Erythematosus of Iraqi Female PatientsGuneyden GuneydenNo ratings yet
- Review Article: Free Radical Biology & Medicine, Vol. 13, Pp. 651-657, 1992Document7 pagesReview Article: Free Radical Biology & Medicine, Vol. 13, Pp. 651-657, 1992MarliCurcioNo ratings yet
- When HeterophyleDocument3 pagesWhen HeterophyleYENY LIZBETH PEREZ HUILLCANo ratings yet
- 10.1515 - CCLM 2023 0825Document6 pages10.1515 - CCLM 2023 0825Arturo Eduardo Huarcaya OntiverosNo ratings yet
- Biomarkers of ImmunosuppressionDocument10 pagesBiomarkers of Immunosuppressionluisc9606No ratings yet
- Behçet Syndrome: From Pathogenesis To Novel Therapies: Clinical and Experimental Medicine December 2014Document13 pagesBehçet Syndrome: From Pathogenesis To Novel Therapies: Clinical and Experimental Medicine December 2014Izamar RomeroNo ratings yet
- HHS Public Access: B Cell Targeted Therapies in Autoimmune DiseaseDocument15 pagesHHS Public Access: B Cell Targeted Therapies in Autoimmune DiseaseLestiNo ratings yet
- Artigo 2Document7 pagesArtigo 2seavidafosseNo ratings yet
- Child Attitude Joward Jllness ScaleDocument1 pageChild Attitude Joward Jllness ScaleAvinash ToraneNo ratings yet
- Systemic Lupus Erythematosus-.-An Autoimmune Disease?: HepatitisDocument6 pagesSystemic Lupus Erythematosus-.-An Autoimmune Disease?: HepatitisDiwa KaranNo ratings yet
- Clinical Immunology Communications: Sandy Nasr, Sara Khalil, Bernard J. Poiesz, Katalin Banki, Andras PerlDocument3 pagesClinical Immunology Communications: Sandy Nasr, Sara Khalil, Bernard J. Poiesz, Katalin Banki, Andras PerlAmarpreet Singh MalhanNo ratings yet
- Fimmu 13 1118180Document4 pagesFimmu 13 1118180Jesus Guadalupe Sanchez MontoyaNo ratings yet
- Blood Group Incompatibility PDFDocument7 pagesBlood Group Incompatibility PDFSekar MarzukiNo ratings yet
- Infection-Genetics Relationship in Systemic Lupus ErythematosusDocument8 pagesInfection-Genetics Relationship in Systemic Lupus ErythematosusegonzalesrnNo ratings yet
- Ankylosing SpondylitisDocument14 pagesAnkylosing SpondylitisJoao MinhoNo ratings yet
- Articulo 3Document14 pagesArticulo 3JL MaravillasNo ratings yet
- Journal of AutoimmunityDocument12 pagesJournal of Autoimmunity2456708893No ratings yet
- Journal Pre-Proof: Cell ReportsDocument57 pagesJournal Pre-Proof: Cell ReportsMohammed Shuaib AhmedNo ratings yet
- Burnett 2019Document15 pagesBurnett 2019diaa skamNo ratings yet
- Microbiome in Sjögren's Syndrome - Here We AreDocument2 pagesMicrobiome in Sjögren's Syndrome - Here We AreMartinaNo ratings yet
- Biomarcadores en InmunosupresionDocument10 pagesBiomarcadores en InmunosupresionRicardo GarciaNo ratings yet
- 1 s2.0 S0753332220305308 MainDocument4 pages1 s2.0 S0753332220305308 Mainvictor rodriguezNo ratings yet
- Hiv 2Document11 pagesHiv 2vn6zmdjgs7No ratings yet
- JRD 30 4 220Document14 pagesJRD 30 4 220Muhammad RijalNo ratings yet
- HIV/AIDS: Immunochemistry, Reductionism and Vaccine Design: A Review of 20 Years of ResearchFrom EverandHIV/AIDS: Immunochemistry, Reductionism and Vaccine Design: A Review of 20 Years of ResearchNo ratings yet
- HLA-C An Accomplice in Rheumatic DiseasesDocument9 pagesHLA-C An Accomplice in Rheumatic DiseasesNaziBrola TsivadzeNo ratings yet
- Correlation Between HLA Haplotypes and TheDocument5 pagesCorrelation Between HLA Haplotypes and TheNaziBrola TsivadzeNo ratings yet
- 12Document4 pages12NaziBrola TsivadzeNo ratings yet
- 24022021144314 - კაციტაძე, ადამიანის ანატომია 2017 1 ტ.Document542 pages24022021144314 - კაციტაძე, ადამიანის ანატომია 2017 1 ტ.NaziBrola TsivadzeNo ratings yet
- 2 Moduli FULL-2225 - Basic LearningDocument27 pages2 Moduli FULL-2225 - Basic LearningNaziBrola TsivadzeNo ratings yet
- Reactive Arthritis: Andres Quiceno, MDDocument16 pagesReactive Arthritis: Andres Quiceno, MDNaziBrola TsivadzeNo ratings yet
- Joint Pain Case StudiesDocument51 pagesJoint Pain Case StudiesNaziBrola TsivadzeNo ratings yet
- Rheumatology Cases For The Internist: Marc C. Hochberg, MD, MPHDocument40 pagesRheumatology Cases For The Internist: Marc C. Hochberg, MD, MPHNaziBrola TsivadzeNo ratings yet
- 1590059744phys.1 CNS Zogadilikv - EEGphys.1 CNS Zogadilikv - EEGDocument33 pages1590059744phys.1 CNS Zogadilikv - EEGphys.1 CNS Zogadilikv - EEGNaziBrola TsivadzeNo ratings yet
- Charlton 2019Document14 pagesCharlton 2019Kikin RizkynnisaNo ratings yet
- Journal Reading - Dr. Sri Rejeki SinagaDocument10 pagesJournal Reading - Dr. Sri Rejeki Sinagaamka chanNo ratings yet
- Acute Nursing Care of The Older Adult With Fragility Hip Fracture: An International Perspective (Part 2)Document26 pagesAcute Nursing Care of The Older Adult With Fragility Hip Fracture: An International Perspective (Part 2)spring flowerNo ratings yet
- Hygiene PDFDocument156 pagesHygiene PDFBob JensonNo ratings yet
- Rheumatic Heart DiseaseDocument7 pagesRheumatic Heart DiseaseRoel John Atamosa CasilacNo ratings yet
- Clinical Considerations Guide - ALLDocument4 pagesClinical Considerations Guide - ALLnenitaNo ratings yet
- Environmental CleaningDocument55 pagesEnvironmental Cleaningriffat shaheenNo ratings yet
- The Effects of Bobath-Based Trunk Exercises On Trunk Control, Functional Capacity, Balance, and Gait: A Pilot Randomized Controlled TrialDocument9 pagesThe Effects of Bobath-Based Trunk Exercises On Trunk Control, Functional Capacity, Balance, and Gait: A Pilot Randomized Controlled Trialnandhini raguNo ratings yet
- Physical Examination 2003Document39 pagesPhysical Examination 2003Gayatri MudliyarNo ratings yet
- Article Information: A B C D eDocument9 pagesArticle Information: A B C D eShifa Auliaputri077No ratings yet
- Health, Safety and Enviironment at ShipyardDocument45 pagesHealth, Safety and Enviironment at ShipyardNaveen DalaniduNo ratings yet
- Antiparasitic Treatment Using Herbs and SpicesDocument11 pagesAntiparasitic Treatment Using Herbs and Spicesrazvan rotaruNo ratings yet
- Medical ReportsDocument1 pageMedical Reportsraheemtimo1No ratings yet
- Blood Bank ProceduresDocument102 pagesBlood Bank Proceduresahalimyousef70% (10)
- Ads 777 Pro Digital Aura Scanner FinalDocument27 pagesAds 777 Pro Digital Aura Scanner Finalmanojjoshisso70No ratings yet
- 1.2 Medical TerminologyDocument101 pages1.2 Medical TerminologyManjot KaurNo ratings yet
- Sinus Massage By: Sushumna Mahesh What Is Sinus Pain?Document5 pagesSinus Massage By: Sushumna Mahesh What Is Sinus Pain?Julenda NikijuluwNo ratings yet
- Lung Cancer PathophysiologyDocument7 pagesLung Cancer PathophysiologyKristian Karl Bautista Kiw-isNo ratings yet
- DivergentesDocument12 pagesDivergentesA LunaNo ratings yet
- The Importance of Empathy ArticleDocument3 pagesThe Importance of Empathy ArticleslvNo ratings yet
- Appearance and General BehaviorDocument3 pagesAppearance and General BehaviorHardeep KaurNo ratings yet
- Nigam A Et Al. Int Surg J. 2020 MayDocument6 pagesNigam A Et Al. Int Surg J. 2020 MaydrpklalNo ratings yet
- Community DewormingDocument55 pagesCommunity DewormingFebNo ratings yet
- Potential Medicinal Plants For Lymphatic Filariasis: A Review Review ArticleDocument7 pagesPotential Medicinal Plants For Lymphatic Filariasis: A Review Review ArticleBALAJI POTBHARENo ratings yet

































































































Download as pdf or txt
You might also like
- Elite 100 NBME Concepts OutlineDocument158 pagesElite 100 NBME Concepts OutlineDeborah Peters100% (3)
- Chapter 36: Aerosol Drug Therapy Test Bank Multiple ChoiceDocument25 pagesChapter 36: Aerosol Drug Therapy Test Bank Multiple ChoiceKhalid Julkanain50% (2)
- Morphine Mikhail BulgakovDocument23 pagesMorphine Mikhail Bulgakovciuciu.oxanaNo ratings yet
- HBsAg WantaiDocument16 pagesHBsAg WantaiAnnisa UtamiNo ratings yet
- Rheumatology Cases Fall Clinics 2019Document85 pagesRheumatology Cases Fall Clinics 2019NaziBrola TsivadzeNo ratings yet
- Ultimate Stretching GuideDocument18 pagesUltimate Stretching GuideJason Collingburn100% (11)
- Chiropractic Technique Self Adjustment Made Easy PDFDocument40 pagesChiropractic Technique Self Adjustment Made Easy PDFEashwar Prasad Meenakshi100% (3)
- Mcmichael 2002Document6 pagesMcmichael 2002peppa pigNo ratings yet
- Does HLA-B27 Status Influence AnkylosingDocument6 pagesDoes HLA-B27 Status Influence AnkylosingcismarcivNo ratings yet
- A Noncompeting Pair of Human Neutralizing Antibodies Block COVID-19 Virus Binding To Its Receptor ACE2Document6 pagesA Noncompeting Pair of Human Neutralizing Antibodies Block COVID-19 Virus Binding To Its Receptor ACE2Walid BoukhelkhalNo ratings yet
- Reactive Arthritis From Clinical Features To PathoDocument11 pagesReactive Arthritis From Clinical Features To Patho[161]Shuaib AktherNo ratings yet
- Bowness 2015 Hla b27Document22 pagesBowness 2015 Hla b27jezy2014No ratings yet
- HLA-B27 Is A Risk Factor For Rheumatoid Arthritis Suggestion ForDocument4 pagesHLA-B27 Is A Risk Factor For Rheumatoid Arthritis Suggestion ForcismarcivNo ratings yet
- COVID-19 Vaccine Design: The Janus Face of Immune EnhancementDocument2 pagesCOVID-19 Vaccine Design: The Janus Face of Immune EnhancementPratiwi PuspanegaraNo ratings yet
- HTTPS:WWW Ncbi NLM Nih gov:pmc:articles:PMC8040313:pdf:ijbsv17p1125Document13 pagesHTTPS:WWW Ncbi NLM Nih gov:pmc:articles:PMC8040313:pdf:ijbsv17p1125Ei DrakorNo ratings yet
- Covid Mda5 Jo1 Mi2Document15 pagesCovid Mda5 Jo1 Mi2Raul ReyesNo ratings yet
- 534 FullDocument9 pages534 FullSoehalino HalNo ratings yet
- Voruganti and Bowness 2020 - New Developments in Our Understanding of Ankylosing Spondylitis Patho-AnnotatedDocument9 pagesVoruganti and Bowness 2020 - New Developments in Our Understanding of Ankylosing Spondylitis Patho-AnnotatedmuamarrayNo ratings yet
- New Pathogenic Hypotheses For SpondyloarthropathiesDocument9 pagesNew Pathogenic Hypotheses For SpondyloarthropathiesAli ChaconNo ratings yet
- Parvovirus Lupus PDFDocument3 pagesParvovirus Lupus PDFDaniel PintoNo ratings yet
- Kel 329Document5 pagesKel 329dr.rahafturkiNo ratings yet
- How To Interpret and Use COVID-19 Serology and Immunology TestsDocument6 pagesHow To Interpret and Use COVID-19 Serology and Immunology TestsDana MNo ratings yet
- Science Abc2241 FullDocument8 pagesScience Abc2241 FullEconomy 365No ratings yet
- ABO Blood Group Type Does Not Influence The LevelDocument8 pagesABO Blood Group Type Does Not Influence The LevelANANo ratings yet
- Bowness 2015Document22 pagesBowness 2015shijumon8055No ratings yet
- Comment: The Potential Danger of Suboptimal Antibody Responses in COVID-19Document3 pagesComment: The Potential Danger of Suboptimal Antibody Responses in COVID-19Flavia Oliveira PampolhaNo ratings yet
- Genetics of Axial SpondyloarthritisDocument1 pageGenetics of Axial SpondyloarthritiscismarcivNo ratings yet
- HLA-B27 What's NewDocument11 pagesHLA-B27 What's NewcismarcivNo ratings yet
- Aps Etiologia PatogenesisDocument21 pagesAps Etiologia PatogenesisLeonardo GarciaNo ratings yet
- Ankylosing Spondylitis HLA-B27 and Klebsiella - AnDocument32 pagesAnkylosing Spondylitis HLA-B27 and Klebsiella - AnbciebienNo ratings yet
- HLA-C An Accomplice in Rheumatic DiseasesDocument9 pagesHLA-C An Accomplice in Rheumatic DiseasesNaziBrola TsivadzeNo ratings yet
- s12026 018 9045 0Document10 pagess12026 018 9045 0ndsjNo ratings yet
- Regular Article: Concurrent in Uenza Vaccination Reduces anti-FVIII Antibody Responses in Murine Hemophilia ADocument11 pagesRegular Article: Concurrent in Uenza Vaccination Reduces anti-FVIII Antibody Responses in Murine Hemophilia Ahasemana hasemanaNo ratings yet
- Review Article: Pathogenic and Epiphenomenal Anti-DNA Antibodies in SLEDocument18 pagesReview Article: Pathogenic and Epiphenomenal Anti-DNA Antibodies in SLEvishnupgiNo ratings yet
- Progress in Guillain Barr Syndrome Immunotherapy A Narrative Review of New Strategies in Recent YearsDocument9 pagesProgress in Guillain Barr Syndrome Immunotherapy A Narrative Review of New Strategies in Recent YearsReem YousifNo ratings yet
- PIIS2589537023003310Document10 pagesPIIS25895370230033109jz5dzj5gwNo ratings yet
- Richardson 2017Document8 pagesRichardson 2017YogiAnantaNo ratings yet
- Cei0121 0270Document5 pagesCei0121 0270Nur WahyuniNo ratings yet
- A Comprehensive Review of Biological Agents For Lupus Beyond Single TargetDocument11 pagesA Comprehensive Review of Biological Agents For Lupus Beyond Single TargetBelen LemaNo ratings yet
- The SARS-Coronavirus-Host Interactome: Identification of Cyclophilins As Target For Pan-Coronavirus InhibitorsDocument15 pagesThe SARS-Coronavirus-Host Interactome: Identification of Cyclophilins As Target For Pan-Coronavirus Inhibitorsfaris daboolNo ratings yet
- Vaccination Versus Hybrid ImmunizationDocument9 pagesVaccination Versus Hybrid ImmunizationJovanaNo ratings yet
- B Cell Responses To HIV InfectionDocument30 pagesB Cell Responses To HIV InfectionLhorruama DiasNo ratings yet
- The Etiological Role of Chlamydia Pneumoniae and Erythematosus of Iraqi Female PatientsDocument5 pagesThe Etiological Role of Chlamydia Pneumoniae and Erythematosus of Iraqi Female PatientsGuneyden GuneydenNo ratings yet
- Review Article: Free Radical Biology & Medicine, Vol. 13, Pp. 651-657, 1992Document7 pagesReview Article: Free Radical Biology & Medicine, Vol. 13, Pp. 651-657, 1992MarliCurcioNo ratings yet
- When HeterophyleDocument3 pagesWhen HeterophyleYENY LIZBETH PEREZ HUILLCANo ratings yet
- 10.1515 - CCLM 2023 0825Document6 pages10.1515 - CCLM 2023 0825Arturo Eduardo Huarcaya OntiverosNo ratings yet
- Biomarkers of ImmunosuppressionDocument10 pagesBiomarkers of Immunosuppressionluisc9606No ratings yet
- Behçet Syndrome: From Pathogenesis To Novel Therapies: Clinical and Experimental Medicine December 2014Document13 pagesBehçet Syndrome: From Pathogenesis To Novel Therapies: Clinical and Experimental Medicine December 2014Izamar RomeroNo ratings yet
- HHS Public Access: B Cell Targeted Therapies in Autoimmune DiseaseDocument15 pagesHHS Public Access: B Cell Targeted Therapies in Autoimmune DiseaseLestiNo ratings yet
- Artigo 2Document7 pagesArtigo 2seavidafosseNo ratings yet
- Child Attitude Joward Jllness ScaleDocument1 pageChild Attitude Joward Jllness ScaleAvinash ToraneNo ratings yet
- Systemic Lupus Erythematosus-.-An Autoimmune Disease?: HepatitisDocument6 pagesSystemic Lupus Erythematosus-.-An Autoimmune Disease?: HepatitisDiwa KaranNo ratings yet
- Clinical Immunology Communications: Sandy Nasr, Sara Khalil, Bernard J. Poiesz, Katalin Banki, Andras PerlDocument3 pagesClinical Immunology Communications: Sandy Nasr, Sara Khalil, Bernard J. Poiesz, Katalin Banki, Andras PerlAmarpreet Singh MalhanNo ratings yet
- Fimmu 13 1118180Document4 pagesFimmu 13 1118180Jesus Guadalupe Sanchez MontoyaNo ratings yet
- Blood Group Incompatibility PDFDocument7 pagesBlood Group Incompatibility PDFSekar MarzukiNo ratings yet
- Infection-Genetics Relationship in Systemic Lupus ErythematosusDocument8 pagesInfection-Genetics Relationship in Systemic Lupus ErythematosusegonzalesrnNo ratings yet
- Ankylosing SpondylitisDocument14 pagesAnkylosing SpondylitisJoao MinhoNo ratings yet
- Articulo 3Document14 pagesArticulo 3JL MaravillasNo ratings yet
- Journal of AutoimmunityDocument12 pagesJournal of Autoimmunity2456708893No ratings yet
- Journal Pre-Proof: Cell ReportsDocument57 pagesJournal Pre-Proof: Cell ReportsMohammed Shuaib AhmedNo ratings yet
- Burnett 2019Document15 pagesBurnett 2019diaa skamNo ratings yet
- Microbiome in Sjögren's Syndrome - Here We AreDocument2 pagesMicrobiome in Sjögren's Syndrome - Here We AreMartinaNo ratings yet
- Biomarcadores en InmunosupresionDocument10 pagesBiomarcadores en InmunosupresionRicardo GarciaNo ratings yet
- 1 s2.0 S0753332220305308 MainDocument4 pages1 s2.0 S0753332220305308 Mainvictor rodriguezNo ratings yet
- Hiv 2Document11 pagesHiv 2vn6zmdjgs7No ratings yet
- JRD 30 4 220Document14 pagesJRD 30 4 220Muhammad RijalNo ratings yet
- HIV/AIDS: Immunochemistry, Reductionism and Vaccine Design: A Review of 20 Years of ResearchFrom EverandHIV/AIDS: Immunochemistry, Reductionism and Vaccine Design: A Review of 20 Years of ResearchNo ratings yet
- HLA-C An Accomplice in Rheumatic DiseasesDocument9 pagesHLA-C An Accomplice in Rheumatic DiseasesNaziBrola TsivadzeNo ratings yet
- Correlation Between HLA Haplotypes and TheDocument5 pagesCorrelation Between HLA Haplotypes and TheNaziBrola TsivadzeNo ratings yet
- 12Document4 pages12NaziBrola TsivadzeNo ratings yet
- 24022021144314 - კაციტაძე, ადამიანის ანატომია 2017 1 ტ.Document542 pages24022021144314 - კაციტაძე, ადამიანის ანატომია 2017 1 ტ.NaziBrola TsivadzeNo ratings yet
- 2 Moduli FULL-2225 - Basic LearningDocument27 pages2 Moduli FULL-2225 - Basic LearningNaziBrola TsivadzeNo ratings yet
- Reactive Arthritis: Andres Quiceno, MDDocument16 pagesReactive Arthritis: Andres Quiceno, MDNaziBrola TsivadzeNo ratings yet
- Joint Pain Case StudiesDocument51 pagesJoint Pain Case StudiesNaziBrola TsivadzeNo ratings yet
- Rheumatology Cases For The Internist: Marc C. Hochberg, MD, MPHDocument40 pagesRheumatology Cases For The Internist: Marc C. Hochberg, MD, MPHNaziBrola TsivadzeNo ratings yet
- 1590059744phys.1 CNS Zogadilikv - EEGphys.1 CNS Zogadilikv - EEGDocument33 pages1590059744phys.1 CNS Zogadilikv - EEGphys.1 CNS Zogadilikv - EEGNaziBrola TsivadzeNo ratings yet
- Charlton 2019Document14 pagesCharlton 2019Kikin RizkynnisaNo ratings yet
- Journal Reading - Dr. Sri Rejeki SinagaDocument10 pagesJournal Reading - Dr. Sri Rejeki Sinagaamka chanNo ratings yet
- Acute Nursing Care of The Older Adult With Fragility Hip Fracture: An International Perspective (Part 2)Document26 pagesAcute Nursing Care of The Older Adult With Fragility Hip Fracture: An International Perspective (Part 2)spring flowerNo ratings yet
- Hygiene PDFDocument156 pagesHygiene PDFBob JensonNo ratings yet
- Rheumatic Heart DiseaseDocument7 pagesRheumatic Heart DiseaseRoel John Atamosa CasilacNo ratings yet
- Clinical Considerations Guide - ALLDocument4 pagesClinical Considerations Guide - ALLnenitaNo ratings yet
- Environmental CleaningDocument55 pagesEnvironmental Cleaningriffat shaheenNo ratings yet
- The Effects of Bobath-Based Trunk Exercises On Trunk Control, Functional Capacity, Balance, and Gait: A Pilot Randomized Controlled TrialDocument9 pagesThe Effects of Bobath-Based Trunk Exercises On Trunk Control, Functional Capacity, Balance, and Gait: A Pilot Randomized Controlled Trialnandhini raguNo ratings yet
- Physical Examination 2003Document39 pagesPhysical Examination 2003Gayatri MudliyarNo ratings yet
- Article Information: A B C D eDocument9 pagesArticle Information: A B C D eShifa Auliaputri077No ratings yet
- Health, Safety and Enviironment at ShipyardDocument45 pagesHealth, Safety and Enviironment at ShipyardNaveen DalaniduNo ratings yet
- Antiparasitic Treatment Using Herbs and SpicesDocument11 pagesAntiparasitic Treatment Using Herbs and Spicesrazvan rotaruNo ratings yet
- Medical ReportsDocument1 pageMedical Reportsraheemtimo1No ratings yet
- Blood Bank ProceduresDocument102 pagesBlood Bank Proceduresahalimyousef70% (10)
- Ads 777 Pro Digital Aura Scanner FinalDocument27 pagesAds 777 Pro Digital Aura Scanner Finalmanojjoshisso70No ratings yet
- 1.2 Medical TerminologyDocument101 pages1.2 Medical TerminologyManjot KaurNo ratings yet
- Sinus Massage By: Sushumna Mahesh What Is Sinus Pain?Document5 pagesSinus Massage By: Sushumna Mahesh What Is Sinus Pain?Julenda NikijuluwNo ratings yet
- Lung Cancer PathophysiologyDocument7 pagesLung Cancer PathophysiologyKristian Karl Bautista Kiw-isNo ratings yet
- DivergentesDocument12 pagesDivergentesA LunaNo ratings yet
- The Importance of Empathy ArticleDocument3 pagesThe Importance of Empathy ArticleslvNo ratings yet
- Appearance and General BehaviorDocument3 pagesAppearance and General BehaviorHardeep KaurNo ratings yet
- Nigam A Et Al. Int Surg J. 2020 MayDocument6 pagesNigam A Et Al. Int Surg J. 2020 MaydrpklalNo ratings yet
- Community DewormingDocument55 pagesCommunity DewormingFebNo ratings yet
- Potential Medicinal Plants For Lymphatic Filariasis: A Review Review ArticleDocument7 pagesPotential Medicinal Plants For Lymphatic Filariasis: A Review Review ArticleBALAJI POTBHARENo ratings yet