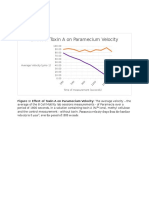
Download as pdf or txt
You might also like
- Biology 1A-Cell Motility Results and ConclusionsDocument6 pagesBiology 1A-Cell Motility Results and ConclusionsAna Maria100% (2)
- Bosniak ClassificationDocument19 pagesBosniak ClassificationGirish Kumar100% (1)
- BBRC Vol 14 No 04 2021-69Document7 pagesBBRC Vol 14 No 04 2021-69Dr Sharique AliNo ratings yet
- Clinical Manifestations, Antimicrobial Resistance and Genomic Feature Analysis of Multidrug-Resistant Elizabethkingia StrainsDocument10 pagesClinical Manifestations, Antimicrobial Resistance and Genomic Feature Analysis of Multidrug-Resistant Elizabethkingia Strainsd4rkgr455No ratings yet
- Anjum Et Al.,2024Document22 pagesAnjum Et Al.,2024Rebeca Garcia RuizNo ratings yet
- Antifungal SusDocument4 pagesAntifungal SusJampala SrinivasNo ratings yet
- Evaluating Molecular Epidemiology of Carbapenem Non-Susceptible Klebsiella Pneumoniae Isolates With MLST, MALDI-TOF MS, PFGEDocument11 pagesEvaluating Molecular Epidemiology of Carbapenem Non-Susceptible Klebsiella Pneumoniae Isolates With MLST, MALDI-TOF MS, PFGEd4rkgr455No ratings yet
- Genotypic Characterization of Mycobacterium Tuberculosis Strains Resistant To Rifampicin Isoniazid and Second Line Antibiotics in ChadDocument14 pagesGenotypic Characterization of Mycobacterium Tuberculosis Strains Resistant To Rifampicin Isoniazid and Second Line Antibiotics in ChadAthenaeum Scientific PublishersNo ratings yet
- Wa0011.Document11 pagesWa0011.umayamajidNo ratings yet
- Antimicrobial Resistance Prediction by Clinical Metagenomics in Pediatric Severe Pneumonia PatientsDocument9 pagesAntimicrobial Resistance Prediction by Clinical Metagenomics in Pediatric Severe Pneumonia Patientsd4rkgr455No ratings yet
- Lembar Jawaban SoalDocument5 pagesLembar Jawaban SoalWahyuNo ratings yet
- Development of A Novel Loop-Mediated Isothermal Amplification Method For The Rapid Detection of Monkeypox Virus InfectionsDocument12 pagesDevelopment of A Novel Loop-Mediated Isothermal Amplification Method For The Rapid Detection of Monkeypox Virus InfectionsV K kasaragodNo ratings yet
- MycoDocument10 pagesMycorajaniNo ratings yet
- Genomic Epidemiology Reveals Multiple Mechanisms of Linezolid Resistance in Clinical Enterococci in ChinaDocument11 pagesGenomic Epidemiology Reveals Multiple Mechanisms of Linezolid Resistance in Clinical Enterococci in Chinad4rkgr455No ratings yet
- 08 - Genome Res.-2012-Casali-735-45Document12 pages08 - Genome Res.-2012-Casali-735-45denisfr79No ratings yet
- ASMNGS 2018 AbstractsDocument187 pagesASMNGS 2018 AbstractsturnersdNo ratings yet
- Antibiotics 10 00419 v2Document19 pagesAntibiotics 10 00419 v2Nora LeeNo ratings yet
- Engineering Coronaviruses To Evaluate Emergence and Pathogenic Potential - Trends in MicrobiologyDocument6 pagesEngineering Coronaviruses To Evaluate Emergence and Pathogenic Potential - Trends in MicrobiologyZsi GaNo ratings yet
- Jci 133 173156Document11 pagesJci 133 173156Dhivya l.sNo ratings yet
- Association of Virulence Genes With Antibiotic Resistance in Pakistani Uropathogenic E. Coli IsolatesDocument6 pagesAssociation of Virulence Genes With Antibiotic Resistance in Pakistani Uropathogenic E. Coli Isolateskhurry testNo ratings yet
- Clinical Characteristics and Drug Resistance of Nocardia in Henan, China, 2017-2023Document14 pagesClinical Characteristics and Drug Resistance of Nocardia in Henan, China, 2017-2023d4rkgr455No ratings yet
- Intestinal Colonization With Multidrug-Resistant Enterobacteriaceae in A Healthy Adult PopulationDocument8 pagesIntestinal Colonization With Multidrug-Resistant Enterobacteriaceae in A Healthy Adult PopulationOpenaccess Research paperNo ratings yet
- Molecular Characterization of Extensively Drug-Resistant Hypervirulent Pseudomonas Aeruginosa Isolates in ChinaDocument9 pagesMolecular Characterization of Extensively Drug-Resistant Hypervirulent Pseudomonas Aeruginosa Isolates in Chinad4rkgr455No ratings yet
- 2020 - Book - MERSCoronavirus LIBRO PDFDocument226 pages2020 - Book - MERSCoronavirus LIBRO PDFJhoel Sebastian Torres GaonaNo ratings yet
- Ajmb2024142 31070522Document18 pagesAjmb2024142 31070522Sakura TraduçõesNo ratings yet
- Microbiolspec - Bai 0021 2019Document11 pagesMicrobiolspec - Bai 0021 2019Franck AMEGNAGLONo ratings yet
- Antibiotic Susceptibility Patterns of Methicillin Resistant Staphyl - PDF NIHDocument5 pagesAntibiotic Susceptibility Patterns of Methicillin Resistant Staphyl - PDF NIHzia ul RahmanNo ratings yet
- Fmicb 13 1073057Document14 pagesFmicb 13 1073057HoàngMinhNguyễnNo ratings yet
- spectrum.00314-23Document12 pagesspectrum.00314-23Muchamad DafipNo ratings yet
- Phenotypic and Genotypic Analysis of Antimicrobial Resistance and Population Structure of Gastroenteritis-Related Aeromonas IsolatesDocument8 pagesPhenotypic and Genotypic Analysis of Antimicrobial Resistance and Population Structure of Gastroenteritis-Related Aeromonas Isolatesd4rkgr455No ratings yet
- 1 s2.0 S1472648320303187 MainDocument17 pages1 s2.0 S1472648320303187 Mainvenny2 tlm polbantenNo ratings yet
- Prevalence of Multidrug Resistance in Streptococcus Pyogenes Isolates From Different Clinical Sources in Al-Diwaniyah CityDocument9 pagesPrevalence of Multidrug Resistance in Streptococcus Pyogenes Isolates From Different Clinical Sources in Al-Diwaniyah CityCentral Asian StudiesNo ratings yet
- 2 PMID 37992081 BiofilmDocument15 pages2 PMID 37992081 Biofilmajaya basnetNo ratings yet
- Antibiotic Resistance Pattern of MRSA Isolates From Pediatric PatientsDocument12 pagesAntibiotic Resistance Pattern of MRSA Isolates From Pediatric PatientsUni NadhilaNo ratings yet
- Etude Maroc CovidDocument25 pagesEtude Maroc CovidAli AmarNo ratings yet
- Klebsiella Pneumoniae Isolated From Urine ofDocument9 pagesKlebsiella Pneumoniae Isolated From Urine ofInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- Latest Advances in Arbovirus DiagnosticsDocument20 pagesLatest Advances in Arbovirus DiagnosticsVictor CostaNo ratings yet
- Bacterial Meningitis in Children Under 15 Years of Age in NepalDocument7 pagesBacterial Meningitis in Children Under 15 Years of Age in NepalPrima YosiNo ratings yet
- Bacterial Contamination On Used Face Masks in Healthcare PersonnelDocument2 pagesBacterial Contamination On Used Face Masks in Healthcare PersonnelMaryam birmahNo ratings yet
- Manuscript Info: International Journal of Advanced ResearchDocument7 pagesManuscript Info: International Journal of Advanced ResearchFajar ReynaldiNo ratings yet
- Genoma TBDocument14 pagesGenoma TBMARIO CASTRONo ratings yet
- KCP 1Document7 pagesKCP 1FurqanRidhaNo ratings yet
- Betteridge Environmentaldrugresistance April 2013Document6 pagesBetteridge Environmentaldrugresistance April 2013Victor GodwinNo ratings yet
- Project Thesis Ready PDFDocument37 pagesProject Thesis Ready PDFAiswarya V ANo ratings yet
- Cto 1545Document10 pagesCto 1545UMT JournalsNo ratings yet
- A Critical Role of Outer Membrane Vesicles in Antibiotic Resistance in Carbapenem-Resistant Klebsiella PneumoniaeDocument12 pagesA Critical Role of Outer Membrane Vesicles in Antibiotic Resistance in Carbapenem-Resistant Klebsiella Pneumoniaed4rkgr455No ratings yet
- Xin 2016, Pfge&MlstDocument8 pagesXin 2016, Pfge&MlstSarah KKCNo ratings yet
- ISSN: 2616 - 0668: UjmrDocument8 pagesISSN: 2616 - 0668: UjmrUMYU Journal of Microbiology Research (UJMR)No ratings yet
- Characterization of Resistance Genes and Replicon Typing in Carbapenem-Resistant Klebsiella Pneumoniae StrainsDocument9 pagesCharacterization of Resistance Genes and Replicon Typing in Carbapenem-Resistant Klebsiella Pneumoniae Strainsd4rkgr455No ratings yet
- 1 PMID 38167427 Antimicrobial ResistancesDocument8 pages1 PMID 38167427 Antimicrobial Resistancesajaya basnetNo ratings yet
- Design and Validation of A Novel Multi-Epitopes Vaccine Against HantavirusDocument12 pagesDesign and Validation of A Novel Multi-Epitopes Vaccine Against HantavirusOgunleke AdebolaNo ratings yet
- Epidemiology IIIDocument7 pagesEpidemiology IIIpaintsilsamuel10No ratings yet
- Large Scale Genomic Analysis of 3067 Sars-Cov-2 Genomes Reveals A Clonal Geo - Distribution and A Rich Genetic Variations of Hotspots MutationsDocument18 pagesLarge Scale Genomic Analysis of 3067 Sars-Cov-2 Genomes Reveals A Clonal Geo - Distribution and A Rich Genetic Variations of Hotspots MutationsSimón Villegas VelásquezNo ratings yet
- Fmicb 06 00791 PDFDocument16 pagesFmicb 06 00791 PDFJhon AraujoNo ratings yet
- 4 THDocument7 pages4 THFatmaNo ratings yet
- Prevalence of Multidrug Resistance in Enterococci Isolates From Different Clinical Sources in Al-Diwaniyah CityDocument9 pagesPrevalence of Multidrug Resistance in Enterococci Isolates From Different Clinical Sources in Al-Diwaniyah CityCentral Asian StudiesNo ratings yet
- International Journal of Infectious DiseasesDocument8 pagesInternational Journal of Infectious DiseasesTiti IndasariNo ratings yet
- Microbiological Indicators of Patients With Confirmed Sars-Cov-2 - InfectionDocument6 pagesMicrobiological Indicators of Patients With Confirmed Sars-Cov-2 - InfectionCentral Asian StudiesNo ratings yet
- 16 4170okeDocument4 pages16 4170okeFenni OktoberryNo ratings yet
- Genomic diversity of β-lactamase producing Pseudomonas aeruginosa in Iran; the impact of global high-risk clonesDocument8 pagesGenomic diversity of β-lactamase producing Pseudomonas aeruginosa in Iran; the impact of global high-risk clonesd4rkgr455No ratings yet
- MedComm Future Medicine - 2024 - Jaishwal - Lessons from COVID‐19 Aspects of prevention therapeutics and diagnosticsDocument32 pagesMedComm Future Medicine - 2024 - Jaishwal - Lessons from COVID‐19 Aspects of prevention therapeutics and diagnosticssprakashsinghNo ratings yet
- Bio 10 Master Study Guide Part 1Document9 pagesBio 10 Master Study Guide Part 1leonor.estimaNo ratings yet
- Sci HubDocument5 pagesSci HubThị Hương NguyễnNo ratings yet
- File 1434571027Document1 pageFile 1434571027Guneyden GuneydenNo ratings yet
- Ejercicios 1Document8 pagesEjercicios 1cokidethNo ratings yet
- PDF Diagnosis and Management of Mitochondrial Disorders Michelangelo Mancuso Ebook Full ChapterDocument53 pagesPDF Diagnosis and Management of Mitochondrial Disorders Michelangelo Mancuso Ebook Full Chapteredward.goodwin495100% (4)
- Pediatric SMLE NotesDocument25 pagesPediatric SMLE NotesMarwa Tariq Ahmed Abdulla Ahmed Al MurbatiNo ratings yet
- Mycoplasmas and Cell-Wall Defective BacteriaDocument26 pagesMycoplasmas and Cell-Wall Defective Bacteriaolivia1026No ratings yet
- Detection of Malarial Parasite in Blood Using Image ProcessingDocument3 pagesDetection of Malarial Parasite in Blood Using Image ProcessingelectricalengineerNo ratings yet
- Open Book Quiz 1 Bit1026 FNH Arg Feb 2018Document4 pagesOpen Book Quiz 1 Bit1026 FNH Arg Feb 2018shubhamNo ratings yet
- Nls Approach in Cancer DiagnosticDocument30 pagesNls Approach in Cancer Diagnosticabap2814No ratings yet
- Rwanda Pricelist-11-11-2016Document85 pagesRwanda Pricelist-11-11-2016BISHWEKA CherifNo ratings yet
- Bacterial Toxins and Its ApplicationDocument10 pagesBacterial Toxins and Its Applicationlamps42No ratings yet
- Poliomyelitis PresentationDocument30 pagesPoliomyelitis PresentationJonah GrantNo ratings yet
- Myasthenia GravisDocument17 pagesMyasthenia GravisjinlaksicNo ratings yet
- Theriogenology I For DVMDocument485 pagesTheriogenology I For DVMFikade AbateNo ratings yet
- 21 382 ImmunologyDocument142 pages21 382 ImmunologyAmirabbas SaffariNo ratings yet
- An Update On Therapeutic Repurposing Strategies For COVID-19Document7 pagesAn Update On Therapeutic Repurposing Strategies For COVID-19Prakash KatakamNo ratings yet
- Isolation of Genomic DNADocument16 pagesIsolation of Genomic DNASamra KousarNo ratings yet
- Human Immunodeficiency Virus Infection / Acquired Immunodeficiency Syndrome (HIV/AIDS) Is A Disease of TheDocument6 pagesHuman Immunodeficiency Virus Infection / Acquired Immunodeficiency Syndrome (HIV/AIDS) Is A Disease of TheDhea Fajeria FitriNo ratings yet
- Biology Unit 3 Revision Year 8Document3 pagesBiology Unit 3 Revision Year 8Alan klnNo ratings yet
- Listening Sample Test 1 Audio ScriptDocument11 pagesListening Sample Test 1 Audio ScriptCCCAXANo ratings yet
- Japanese EncephalitisDocument7 pagesJapanese EncephalitiskafhcompNo ratings yet
- International Biology Olympiad - Hong Kong Contest 2018 - 2018Document70 pagesInternational Biology Olympiad - Hong Kong Contest 2018 - 2018Ngọc CaiNo ratings yet
- GeneDx Variant Classification Process June 2021Document4 pagesGeneDx Variant Classification Process June 2021Liza HavengaNo ratings yet
- PulpaDocument173 pagesPulpaYoshe Kartika SentosaNo ratings yet
- Treatment of Latent Tuberculosis InfectionDocument10 pagesTreatment of Latent Tuberculosis InfectionRichard WooliteNo ratings yet
- Body Condition Scoring of Dairy Cattle A Review - 2 PDFDocument8 pagesBody Condition Scoring of Dairy Cattle A Review - 2 PDFfrankyNo ratings yet
- s41598 018 35903 5Document9 pagess41598 018 35903 5leartaNo ratings yet