
Pulmón e Isquemia Intestinal
Pulmón e Isquemia Intestinal
You might also like
- Differential Effects On Innate Versus Adaptive Immune Responses by WF10Document10 pagesDifferential Effects On Innate Versus Adaptive Immune Responses by WF10Anonymous KUimpSvdNo ratings yet
- Artículo+Document10 pagesArtículo+king130No ratings yet
- Reactive Oxygen Species (ROS) Mediates The Mitochondrial-Dependent Apoptosis Induced by Transforming Growth Factor B in Fetal HepatocytesDocument11 pagesReactive Oxygen Species (ROS) Mediates The Mitochondrial-Dependent Apoptosis Induced by Transforming Growth Factor B in Fetal HepatocytesSameer PashaNo ratings yet
- Expression of A Gene Encoding Trypanosoma Congolense Putative Abc1 Family Protein Is Developmentally RegulatedDocument0 pagesExpression of A Gene Encoding Trypanosoma Congolense Putative Abc1 Family Protein Is Developmentally RegulatedMaísa FonsecaNo ratings yet
- Biomed ..Document10 pagesBiomed ..Romina Molette GonzalesNo ratings yet
- In Vitro Inhibitory Effects of Antioxidants On Cytotoxicity of T-2 ToxinDocument6 pagesIn Vitro Inhibitory Effects of Antioxidants On Cytotoxicity of T-2 ToxinElijah James LegaspiNo ratings yet
- Map KinaseDocument8 pagesMap KinasemoabdelhameidNo ratings yet
- TMP CE4Document12 pagesTMP CE4FrontiersNo ratings yet
- Overexpression of Hematopoietically Expressed Homeoprotein Induces Nonapoptotic Cell Death in Mouse Prechondrogenic ATDC5 CellsDocument7 pagesOverexpression of Hematopoietically Expressed Homeoprotein Induces Nonapoptotic Cell Death in Mouse Prechondrogenic ATDC5 CellsCamilo CabreraNo ratings yet
- Aeruginosa Strain PAO1 Is Regulated by Quorum Sensing andDocument15 pagesAeruginosa Strain PAO1 Is Regulated by Quorum Sensing andSarah KKCNo ratings yet
- 1 s2.0 S0753332218335388 MainDocument8 pages1 s2.0 S0753332218335388 MainReza SNo ratings yet
- Sulforaphane Attenuates Pulmonary Fibrosis by Inhibiting The Epithelial-Mesenchymal TransitionDocument10 pagesSulforaphane Attenuates Pulmonary Fibrosis by Inhibiting The Epithelial-Mesenchymal TransitiongardamdNo ratings yet
- Necrotic Cell Death in Response To Oxidant Stress Involves The Activation of The Apoptogenic Caspase-8 Bid PathwayDocument8 pagesNecrotic Cell Death in Response To Oxidant Stress Involves The Activation of The Apoptogenic Caspase-8 Bid PathwayalihamraghaniNo ratings yet
- Integradora Articulo InglesDocument14 pagesIntegradora Articulo InglesALEJANDRANo ratings yet
- Role of Apoptotic and Necrotic Cell Death Under Physiologic ConditionsDocument10 pagesRole of Apoptotic and Necrotic Cell Death Under Physiologic ConditionsAsish GeiorgeNo ratings yet
- Gentamicin-Induced Apoptosis in Renal Cell Lines and Embryonic Rat FibroblastsDocument0 pagesGentamicin-Induced Apoptosis in Renal Cell Lines and Embryonic Rat FibroblastsCLPHtheoryNo ratings yet
- 58 19Document10 pages58 19Alberto RamirezNo ratings yet
- Exosomas Inmuno1Document10 pagesExosomas Inmuno1Juan GomezNo ratings yet
- AKT/Protein Kinase B Regulation of BCL Family Members During Oxysterol-Induced ApoptosisDocument8 pagesAKT/Protein Kinase B Regulation of BCL Family Members During Oxysterol-Induced ApoptosisNareshNo ratings yet
- Acute HepatotoxityDocument11 pagesAcute HepatotoxityThially BragaNo ratings yet
- 1 s2.0 S0006295221003038 MainDocument14 pages1 s2.0 S0006295221003038 MainDIEGO ARMANDO SARMIENTO BUITRAGONo ratings yet
- Journal ParasitDocument19 pagesJournal ParasitNimade IndriNo ratings yet
- Fpi en Biologia MolecularDocument9 pagesFpi en Biologia Molecularsofia barraganNo ratings yet
- Jun Zou, Nathan ShankarDocument11 pagesJun Zou, Nathan ShankarHQ HQNo ratings yet
- JCI37948 v2Document14 pagesJCI37948 v2AJ FabrizioNo ratings yet
- Hyperoxia in The Intensive Care Unit: Why More Is Not Always BetterDocument6 pagesHyperoxia in The Intensive Care Unit: Why More Is Not Always Betteralbimar239512No ratings yet
- 1953 FullDocument10 pages1953 FullboboyoyotNo ratings yet
- Alkaloids Induce Programmed Cell Death in Bloodstream Forms of Trypanosomes (Trypanosoma B. Brucei)Document12 pagesAlkaloids Induce Programmed Cell Death in Bloodstream Forms of Trypanosomes (Trypanosoma B. Brucei)Kayo PaivaNo ratings yet
- Delfinidina PaperDocument13 pagesDelfinidina PaperCristobal ParraNo ratings yet
- Ayissiowona2013 JQ1Document7 pagesAyissiowona2013 JQ1iforaNo ratings yet
- Biology: Programmed Cell Death 1 Ligand (PD-L1) On T Cells Generates Treg Suppression From MemoryDocument4 pagesBiology: Programmed Cell Death 1 Ligand (PD-L1) On T Cells Generates Treg Suppression From MemoryMarlyNationTVNo ratings yet
- STAT3 Regulates Cytokine-Mediated Generation of Inflammatory Helper T CellsDocument6 pagesSTAT3 Regulates Cytokine-Mediated Generation of Inflammatory Helper T Cellssa571922No ratings yet
- Emetine Produce Entamoeba Histolytica Death by IndDocument6 pagesEmetine Produce Entamoeba Histolytica Death by IndJota CarlosNo ratings yet
- Chen 2009Document11 pagesChen 2009darksoul4040940No ratings yet
- Acute Phase Proteins 1-S2.0-S1090023303001199-MainDocument13 pagesAcute Phase Proteins 1-S2.0-S1090023303001199-MainDirección Científica Laboratorio VitalabNo ratings yet
- II 06 Hernandez P Internat Immunopharmacol. AICD Linfo T and Vimang.2006Document10 pagesII 06 Hernandez P Internat Immunopharmacol. AICD Linfo T and Vimang.2006Mariela Guevara GarcíaNo ratings yet
- 35-Dicaffeoylquinic Acid Protects H9C2 Cells AgainDocument8 pages35-Dicaffeoylquinic Acid Protects H9C2 Cells AgainRekod Tita N11No ratings yet
- Apoptosis: Jalal Ghasemzadeh Andrology LabDocument67 pagesApoptosis: Jalal Ghasemzadeh Andrology LabValeria DezaNo ratings yet
- Digallate (Tf3) : Inhibition of Sars-Cov 3C-Like Protease Activity by Theaflavin-3,3Document8 pagesDigallate (Tf3) : Inhibition of Sars-Cov 3C-Like Protease Activity by Theaflavin-3,3Yovan BlancoNo ratings yet
- Belas Figuras Nada de CampyDocument6 pagesBelas Figuras Nada de CampyHaurusNo ratings yet
- Cao 2015Document10 pagesCao 2015joselouisngNo ratings yet
- Infect. Immun.-1996-Dahl-399-405Document7 pagesInfect. Immun.-1996-Dahl-399-405Nika AndrianiNo ratings yet
- Carocin 2Document12 pagesCarocin 2Leticia AvelarNo ratings yet
- tmp337B TMPDocument29 pagestmp337B TMPFrontiersNo ratings yet
- Phosphatidylinositol 3 - Kinase Signalling Supports Cell Height in Established Epithelial MonolayersDocument11 pagesPhosphatidylinositol 3 - Kinase Signalling Supports Cell Height in Established Epithelial MonolayersMohammed Elias AlamNo ratings yet
- Refolding and Characterization of RecombDocument6 pagesRefolding and Characterization of RecombMobarakNo ratings yet
- Gene Encoding Isopentenyl Diphosphate Isomerase: Escherichia Coli Open Reading Frame 696 Is Idi, A NonessentialDocument6 pagesGene Encoding Isopentenyl Diphosphate Isomerase: Escherichia Coli Open Reading Frame 696 Is Idi, A NonessentialHuong TranNo ratings yet
- Prolactin Increase Frecuency of T Helper FolicularDocument15 pagesProlactin Increase Frecuency of T Helper Folicularauroragamez2015No ratings yet
- 1990 - Recombinant Human Interferon-Y - Differences in Glycosylation and Proteolytic Processing Lead To Heterogeneity in Batch CultureDocument5 pages1990 - Recombinant Human Interferon-Y - Differences in Glycosylation and Proteolytic Processing Lead To Heterogeneity in Batch Culture顏世隆No ratings yet
- Ghetie 1996Document7 pagesGhetie 1996Arjun SNo ratings yet
- J. Biol. Chem.-2015-Manne-11557-68Document13 pagesJ. Biol. Chem.-2015-Manne-11557-68zsyedphd1No ratings yet
- 14th ICI Abstract BookDocument4 pages14th ICI Abstract BookMuhammadGagasSasongkoNo ratings yet
- Protective Roles of Mast Cells Against Enterobacterial Infection Are Mediated by Toll-Like Receptor 4Document7 pagesProtective Roles of Mast Cells Against Enterobacterial Infection Are Mediated by Toll-Like Receptor 4harumiaikoNo ratings yet
- Mitake Mushroom 3Document6 pagesMitake Mushroom 3dayra maría ixa marín salamancaNo ratings yet
- FB2006A0006Document11 pagesFB2006A0006Machlery KunNo ratings yet
- Ijmm 25 4 593 PDFDocument8 pagesIjmm 25 4 593 PDFNickNo ratings yet
- Co-signal Molecules in T Cell Activation: Immune Regulation in Health and DiseaseFrom EverandCo-signal Molecules in T Cell Activation: Immune Regulation in Health and DiseaseMiyuki AzumaNo ratings yet
- Transfer Factors: Properties, Mechanism of Action and Its Clinical ApplicationsFrom EverandTransfer Factors: Properties, Mechanism of Action and Its Clinical ApplicationsNo ratings yet
- Fast Facts: Asthma: Improve patient self-management and drug use, achieve asthma controlFrom EverandFast Facts: Asthma: Improve patient self-management and drug use, achieve asthma controlNo ratings yet
- Luxeon 7070Document22 pagesLuxeon 7070guciNo ratings yet
- Section: Expenditure Subsystem Subsection: 060-01 Subsection: Overview Page 1 of 2Document2 pagesSection: Expenditure Subsystem Subsection: 060-01 Subsection: Overview Page 1 of 2Clea Marie MissionNo ratings yet
- General Ledger Interview Questions in R12Document9 pagesGeneral Ledger Interview Questions in R12jeedNo ratings yet
- Batch Process in Fabrics - Recherche GoogleDocument1 pageBatch Process in Fabrics - Recherche GoogleRamel FalcaoNo ratings yet
- Classification of Imbalanced Malaria Disease UsingDocument5 pagesClassification of Imbalanced Malaria Disease Usinglubna gemaaNo ratings yet
- Day 1 Vastu ShastrahastraDocument16 pagesDay 1 Vastu ShastrahastraPooja BhatiNo ratings yet
- Neonatal JaundiceDocument15 pagesNeonatal JaundiceSharan SekhonNo ratings yet
- Fundamentals of Digital IC Design: ECE 428 Programmable ASIC DesignDocument29 pagesFundamentals of Digital IC Design: ECE 428 Programmable ASIC DesignBadangNo ratings yet
- Circular Flow LessonDocument13 pagesCircular Flow LessonKimball SarmientoNo ratings yet
- Design Calculation of FrameDocument5 pagesDesign Calculation of FrameUdhasu NayakNo ratings yet
- Booklist Grade 3Document3 pagesBooklist Grade 3solid_impactNo ratings yet
- Bridge Course Unit 1 AssignmentDocument2 pagesBridge Course Unit 1 AssignmentSrinidhi SrinivasanNo ratings yet
- Exercise 5: Chapter 6, Section 6.1, Page 396Document2 pagesExercise 5: Chapter 6, Section 6.1, Page 396allauddinbaloch84No ratings yet
- Proposed Solid Sodium Cyanide Plant Upgrade Public Environmental ReviewDocument178 pagesProposed Solid Sodium Cyanide Plant Upgrade Public Environmental ReviewArsalan GhasrsazNo ratings yet
- Contengency PlanDocument10 pagesContengency PlanFernando Dumar Mannod Jr.No ratings yet
- IB Chem, Topic 10, Organic Quiz 1 ReviewDocument4 pagesIB Chem, Topic 10, Organic Quiz 1 ReviewJulie VuNo ratings yet
- Memory Management: Hardware ImplementationDocument11 pagesMemory Management: Hardware ImplementationAashrey JainNo ratings yet
- Zuckerberg Responses To Commerce Committee QFRs1Document229 pagesZuckerberg Responses To Commerce Committee QFRs1TechCrunch100% (2)
- Muar Bio Term 2 TrialDocument12 pagesMuar Bio Term 2 TrialChee Beng YeapNo ratings yet
- Environmental Policy Stringency, Technical Progress and Pollution Haven HypothesisDocument20 pagesEnvironmental Policy Stringency, Technical Progress and Pollution Haven HypothesisNaila ErumNo ratings yet
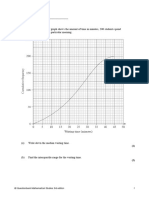
- The Cumulative Frequency Graph Shows The Amount of Time in Minutes, 200 Students Spend Waiting For Their Train On A Particular MorningDocument8 pagesThe Cumulative Frequency Graph Shows The Amount of Time in Minutes, 200 Students Spend Waiting For Their Train On A Particular MorningJonathanNo ratings yet
- Power Engineering PM Check ListDocument11 pagesPower Engineering PM Check ListSelvn SubramanianNo ratings yet
- Sample Specification of FansDocument5 pagesSample Specification of Fanscashlover100% (1)
- Bruce and Youngs Theory of Facial RecognitionDocument2 pagesBruce and Youngs Theory of Facial Recognitionalice100% (1)
- Ang Pag-Aalaga NG Alimango - Beta - 356515 PDFDocument35 pagesAng Pag-Aalaga NG Alimango - Beta - 356515 PDFSamsam WaldNo ratings yet
- Goppinti Alludu FilmDocument3 pagesGoppinti Alludu FilmMadhuswamygaddamNo ratings yet
- Adverse Effects of Hormonal ContraceptivesDocument153 pagesAdverse Effects of Hormonal ContraceptivesTWGAScNo ratings yet
- MN 012479 R02 HV40.000 Chipeo SiteDocument20 pagesMN 012479 R02 HV40.000 Chipeo SiteSecun lasogaNo ratings yet
- 5 Forms of International Business - IbtDocument3 pages5 Forms of International Business - IbtGilbert CastroNo ratings yet
- Introduction To Pharmaceutics.Document10 pagesIntroduction To Pharmaceutics.khelicu517No ratings yet
























































































Download as pdf or txt
You might also like
- Differential Effects On Innate Versus Adaptive Immune Responses by WF10Document10 pagesDifferential Effects On Innate Versus Adaptive Immune Responses by WF10Anonymous KUimpSvdNo ratings yet
- Artículo+Document10 pagesArtículo+king130No ratings yet
- Reactive Oxygen Species (ROS) Mediates The Mitochondrial-Dependent Apoptosis Induced by Transforming Growth Factor B in Fetal HepatocytesDocument11 pagesReactive Oxygen Species (ROS) Mediates The Mitochondrial-Dependent Apoptosis Induced by Transforming Growth Factor B in Fetal HepatocytesSameer PashaNo ratings yet
- Expression of A Gene Encoding Trypanosoma Congolense Putative Abc1 Family Protein Is Developmentally RegulatedDocument0 pagesExpression of A Gene Encoding Trypanosoma Congolense Putative Abc1 Family Protein Is Developmentally RegulatedMaísa FonsecaNo ratings yet
- Biomed ..Document10 pagesBiomed ..Romina Molette GonzalesNo ratings yet
- In Vitro Inhibitory Effects of Antioxidants On Cytotoxicity of T-2 ToxinDocument6 pagesIn Vitro Inhibitory Effects of Antioxidants On Cytotoxicity of T-2 ToxinElijah James LegaspiNo ratings yet
- Map KinaseDocument8 pagesMap KinasemoabdelhameidNo ratings yet
- TMP CE4Document12 pagesTMP CE4FrontiersNo ratings yet
- Overexpression of Hematopoietically Expressed Homeoprotein Induces Nonapoptotic Cell Death in Mouse Prechondrogenic ATDC5 CellsDocument7 pagesOverexpression of Hematopoietically Expressed Homeoprotein Induces Nonapoptotic Cell Death in Mouse Prechondrogenic ATDC5 CellsCamilo CabreraNo ratings yet
- Aeruginosa Strain PAO1 Is Regulated by Quorum Sensing andDocument15 pagesAeruginosa Strain PAO1 Is Regulated by Quorum Sensing andSarah KKCNo ratings yet
- 1 s2.0 S0753332218335388 MainDocument8 pages1 s2.0 S0753332218335388 MainReza SNo ratings yet
- Sulforaphane Attenuates Pulmonary Fibrosis by Inhibiting The Epithelial-Mesenchymal TransitionDocument10 pagesSulforaphane Attenuates Pulmonary Fibrosis by Inhibiting The Epithelial-Mesenchymal TransitiongardamdNo ratings yet
- Necrotic Cell Death in Response To Oxidant Stress Involves The Activation of The Apoptogenic Caspase-8 Bid PathwayDocument8 pagesNecrotic Cell Death in Response To Oxidant Stress Involves The Activation of The Apoptogenic Caspase-8 Bid PathwayalihamraghaniNo ratings yet
- Integradora Articulo InglesDocument14 pagesIntegradora Articulo InglesALEJANDRANo ratings yet
- Role of Apoptotic and Necrotic Cell Death Under Physiologic ConditionsDocument10 pagesRole of Apoptotic and Necrotic Cell Death Under Physiologic ConditionsAsish GeiorgeNo ratings yet
- Gentamicin-Induced Apoptosis in Renal Cell Lines and Embryonic Rat FibroblastsDocument0 pagesGentamicin-Induced Apoptosis in Renal Cell Lines and Embryonic Rat FibroblastsCLPHtheoryNo ratings yet
- 58 19Document10 pages58 19Alberto RamirezNo ratings yet
- Exosomas Inmuno1Document10 pagesExosomas Inmuno1Juan GomezNo ratings yet
- AKT/Protein Kinase B Regulation of BCL Family Members During Oxysterol-Induced ApoptosisDocument8 pagesAKT/Protein Kinase B Regulation of BCL Family Members During Oxysterol-Induced ApoptosisNareshNo ratings yet
- Acute HepatotoxityDocument11 pagesAcute HepatotoxityThially BragaNo ratings yet
- 1 s2.0 S0006295221003038 MainDocument14 pages1 s2.0 S0006295221003038 MainDIEGO ARMANDO SARMIENTO BUITRAGONo ratings yet
- Journal ParasitDocument19 pagesJournal ParasitNimade IndriNo ratings yet
- Fpi en Biologia MolecularDocument9 pagesFpi en Biologia Molecularsofia barraganNo ratings yet
- Jun Zou, Nathan ShankarDocument11 pagesJun Zou, Nathan ShankarHQ HQNo ratings yet
- JCI37948 v2Document14 pagesJCI37948 v2AJ FabrizioNo ratings yet
- Hyperoxia in The Intensive Care Unit: Why More Is Not Always BetterDocument6 pagesHyperoxia in The Intensive Care Unit: Why More Is Not Always Betteralbimar239512No ratings yet
- 1953 FullDocument10 pages1953 FullboboyoyotNo ratings yet
- Alkaloids Induce Programmed Cell Death in Bloodstream Forms of Trypanosomes (Trypanosoma B. Brucei)Document12 pagesAlkaloids Induce Programmed Cell Death in Bloodstream Forms of Trypanosomes (Trypanosoma B. Brucei)Kayo PaivaNo ratings yet
- Delfinidina PaperDocument13 pagesDelfinidina PaperCristobal ParraNo ratings yet
- Ayissiowona2013 JQ1Document7 pagesAyissiowona2013 JQ1iforaNo ratings yet
- Biology: Programmed Cell Death 1 Ligand (PD-L1) On T Cells Generates Treg Suppression From MemoryDocument4 pagesBiology: Programmed Cell Death 1 Ligand (PD-L1) On T Cells Generates Treg Suppression From MemoryMarlyNationTVNo ratings yet
- STAT3 Regulates Cytokine-Mediated Generation of Inflammatory Helper T CellsDocument6 pagesSTAT3 Regulates Cytokine-Mediated Generation of Inflammatory Helper T Cellssa571922No ratings yet
- Emetine Produce Entamoeba Histolytica Death by IndDocument6 pagesEmetine Produce Entamoeba Histolytica Death by IndJota CarlosNo ratings yet
- Chen 2009Document11 pagesChen 2009darksoul4040940No ratings yet
- Acute Phase Proteins 1-S2.0-S1090023303001199-MainDocument13 pagesAcute Phase Proteins 1-S2.0-S1090023303001199-MainDirección Científica Laboratorio VitalabNo ratings yet
- II 06 Hernandez P Internat Immunopharmacol. AICD Linfo T and Vimang.2006Document10 pagesII 06 Hernandez P Internat Immunopharmacol. AICD Linfo T and Vimang.2006Mariela Guevara GarcíaNo ratings yet
- 35-Dicaffeoylquinic Acid Protects H9C2 Cells AgainDocument8 pages35-Dicaffeoylquinic Acid Protects H9C2 Cells AgainRekod Tita N11No ratings yet
- Apoptosis: Jalal Ghasemzadeh Andrology LabDocument67 pagesApoptosis: Jalal Ghasemzadeh Andrology LabValeria DezaNo ratings yet
- Digallate (Tf3) : Inhibition of Sars-Cov 3C-Like Protease Activity by Theaflavin-3,3Document8 pagesDigallate (Tf3) : Inhibition of Sars-Cov 3C-Like Protease Activity by Theaflavin-3,3Yovan BlancoNo ratings yet
- Belas Figuras Nada de CampyDocument6 pagesBelas Figuras Nada de CampyHaurusNo ratings yet
- Cao 2015Document10 pagesCao 2015joselouisngNo ratings yet
- Infect. Immun.-1996-Dahl-399-405Document7 pagesInfect. Immun.-1996-Dahl-399-405Nika AndrianiNo ratings yet
- Carocin 2Document12 pagesCarocin 2Leticia AvelarNo ratings yet
- tmp337B TMPDocument29 pagestmp337B TMPFrontiersNo ratings yet
- Phosphatidylinositol 3 - Kinase Signalling Supports Cell Height in Established Epithelial MonolayersDocument11 pagesPhosphatidylinositol 3 - Kinase Signalling Supports Cell Height in Established Epithelial MonolayersMohammed Elias AlamNo ratings yet
- Refolding and Characterization of RecombDocument6 pagesRefolding and Characterization of RecombMobarakNo ratings yet
- Gene Encoding Isopentenyl Diphosphate Isomerase: Escherichia Coli Open Reading Frame 696 Is Idi, A NonessentialDocument6 pagesGene Encoding Isopentenyl Diphosphate Isomerase: Escherichia Coli Open Reading Frame 696 Is Idi, A NonessentialHuong TranNo ratings yet
- Prolactin Increase Frecuency of T Helper FolicularDocument15 pagesProlactin Increase Frecuency of T Helper Folicularauroragamez2015No ratings yet
- 1990 - Recombinant Human Interferon-Y - Differences in Glycosylation and Proteolytic Processing Lead To Heterogeneity in Batch CultureDocument5 pages1990 - Recombinant Human Interferon-Y - Differences in Glycosylation and Proteolytic Processing Lead To Heterogeneity in Batch Culture顏世隆No ratings yet
- Ghetie 1996Document7 pagesGhetie 1996Arjun SNo ratings yet
- J. Biol. Chem.-2015-Manne-11557-68Document13 pagesJ. Biol. Chem.-2015-Manne-11557-68zsyedphd1No ratings yet
- 14th ICI Abstract BookDocument4 pages14th ICI Abstract BookMuhammadGagasSasongkoNo ratings yet
- Protective Roles of Mast Cells Against Enterobacterial Infection Are Mediated by Toll-Like Receptor 4Document7 pagesProtective Roles of Mast Cells Against Enterobacterial Infection Are Mediated by Toll-Like Receptor 4harumiaikoNo ratings yet
- Mitake Mushroom 3Document6 pagesMitake Mushroom 3dayra maría ixa marín salamancaNo ratings yet
- FB2006A0006Document11 pagesFB2006A0006Machlery KunNo ratings yet
- Ijmm 25 4 593 PDFDocument8 pagesIjmm 25 4 593 PDFNickNo ratings yet
- Co-signal Molecules in T Cell Activation: Immune Regulation in Health and DiseaseFrom EverandCo-signal Molecules in T Cell Activation: Immune Regulation in Health and DiseaseMiyuki AzumaNo ratings yet
- Transfer Factors: Properties, Mechanism of Action and Its Clinical ApplicationsFrom EverandTransfer Factors: Properties, Mechanism of Action and Its Clinical ApplicationsNo ratings yet
- Fast Facts: Asthma: Improve patient self-management and drug use, achieve asthma controlFrom EverandFast Facts: Asthma: Improve patient self-management and drug use, achieve asthma controlNo ratings yet
- Luxeon 7070Document22 pagesLuxeon 7070guciNo ratings yet
- Section: Expenditure Subsystem Subsection: 060-01 Subsection: Overview Page 1 of 2Document2 pagesSection: Expenditure Subsystem Subsection: 060-01 Subsection: Overview Page 1 of 2Clea Marie MissionNo ratings yet
- General Ledger Interview Questions in R12Document9 pagesGeneral Ledger Interview Questions in R12jeedNo ratings yet
- Batch Process in Fabrics - Recherche GoogleDocument1 pageBatch Process in Fabrics - Recherche GoogleRamel FalcaoNo ratings yet
- Classification of Imbalanced Malaria Disease UsingDocument5 pagesClassification of Imbalanced Malaria Disease Usinglubna gemaaNo ratings yet
- Day 1 Vastu ShastrahastraDocument16 pagesDay 1 Vastu ShastrahastraPooja BhatiNo ratings yet
- Neonatal JaundiceDocument15 pagesNeonatal JaundiceSharan SekhonNo ratings yet
- Fundamentals of Digital IC Design: ECE 428 Programmable ASIC DesignDocument29 pagesFundamentals of Digital IC Design: ECE 428 Programmable ASIC DesignBadangNo ratings yet
- Circular Flow LessonDocument13 pagesCircular Flow LessonKimball SarmientoNo ratings yet
- Design Calculation of FrameDocument5 pagesDesign Calculation of FrameUdhasu NayakNo ratings yet
- Booklist Grade 3Document3 pagesBooklist Grade 3solid_impactNo ratings yet
- Bridge Course Unit 1 AssignmentDocument2 pagesBridge Course Unit 1 AssignmentSrinidhi SrinivasanNo ratings yet
- Exercise 5: Chapter 6, Section 6.1, Page 396Document2 pagesExercise 5: Chapter 6, Section 6.1, Page 396allauddinbaloch84No ratings yet
- Proposed Solid Sodium Cyanide Plant Upgrade Public Environmental ReviewDocument178 pagesProposed Solid Sodium Cyanide Plant Upgrade Public Environmental ReviewArsalan GhasrsazNo ratings yet
- Contengency PlanDocument10 pagesContengency PlanFernando Dumar Mannod Jr.No ratings yet
- IB Chem, Topic 10, Organic Quiz 1 ReviewDocument4 pagesIB Chem, Topic 10, Organic Quiz 1 ReviewJulie VuNo ratings yet
- Memory Management: Hardware ImplementationDocument11 pagesMemory Management: Hardware ImplementationAashrey JainNo ratings yet
- Zuckerberg Responses To Commerce Committee QFRs1Document229 pagesZuckerberg Responses To Commerce Committee QFRs1TechCrunch100% (2)
- Muar Bio Term 2 TrialDocument12 pagesMuar Bio Term 2 TrialChee Beng YeapNo ratings yet
- Environmental Policy Stringency, Technical Progress and Pollution Haven HypothesisDocument20 pagesEnvironmental Policy Stringency, Technical Progress and Pollution Haven HypothesisNaila ErumNo ratings yet
- The Cumulative Frequency Graph Shows The Amount of Time in Minutes, 200 Students Spend Waiting For Their Train On A Particular MorningDocument8 pagesThe Cumulative Frequency Graph Shows The Amount of Time in Minutes, 200 Students Spend Waiting For Their Train On A Particular MorningJonathanNo ratings yet
- Power Engineering PM Check ListDocument11 pagesPower Engineering PM Check ListSelvn SubramanianNo ratings yet
- Sample Specification of FansDocument5 pagesSample Specification of Fanscashlover100% (1)
- Bruce and Youngs Theory of Facial RecognitionDocument2 pagesBruce and Youngs Theory of Facial Recognitionalice100% (1)
- Ang Pag-Aalaga NG Alimango - Beta - 356515 PDFDocument35 pagesAng Pag-Aalaga NG Alimango - Beta - 356515 PDFSamsam WaldNo ratings yet
- Goppinti Alludu FilmDocument3 pagesGoppinti Alludu FilmMadhuswamygaddamNo ratings yet
- Adverse Effects of Hormonal ContraceptivesDocument153 pagesAdverse Effects of Hormonal ContraceptivesTWGAScNo ratings yet
- MN 012479 R02 HV40.000 Chipeo SiteDocument20 pagesMN 012479 R02 HV40.000 Chipeo SiteSecun lasogaNo ratings yet
- 5 Forms of International Business - IbtDocument3 pages5 Forms of International Business - IbtGilbert CastroNo ratings yet
- Introduction To Pharmaceutics.Document10 pagesIntroduction To Pharmaceutics.khelicu517No ratings yet