
nn2c02714 Si 001
nn2c02714 Si 001
You might also like
- Biology 2nd Lab Report (Onion Cell)Document4 pagesBiology 2nd Lab Report (Onion Cell)Johanna Haludilu89% (18)
- Cells Exploration ActivitiesDocument32 pagesCells Exploration ActivitiesSay KhoNo ratings yet
- Cobas U 701 Operators Manual EN (Initial Version) 1.0.0 ENDocument238 pagesCobas U 701 Operators Manual EN (Initial Version) 1.0.0 ENRicky100% (1)
- Ja4c05178 Si 001Document9 pagesJa4c05178 Si 001Yong-Sheng WeiNo ratings yet
- La0c02115 Si 001Document13 pagesLa0c02115 Si 001peeyush kumarNo ratings yet
- Basis of Computed Radiography & PACS: Digital Film-ScreenDocument28 pagesBasis of Computed Radiography & PACS: Digital Film-ScreenAmar MohammedNo ratings yet
- Dyes SupportDocument7 pagesDyes SupportawaisNo ratings yet
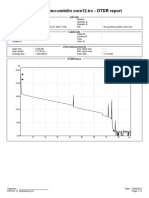
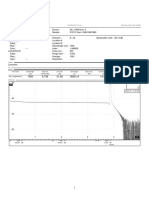
- NTN Pauhlimo-Ombilin Core12Document2 pagesNTN Pauhlimo-Ombilin Core12rahman.ab1212No ratings yet
- Silva 2014CO2解离的微波脉冲放电的光学表征Document14 pagesSilva 2014CO2解离的微波脉冲放电的光学表征yuan MoNo ratings yet
- ImagingDocument29 pagesImagingDyhoiNo ratings yet
- 12mihailidis ElectronBeamDosimetryDocument92 pages12mihailidis ElectronBeamDosimetryitumNo ratings yet
- Tribological Properties of Diamond-Like Carbon Films Produced by Different Deposition TechniquesDocument5 pagesTribological Properties of Diamond-Like Carbon Films Produced by Different Deposition TechniquesEngels OchoaNo ratings yet
- Chapter-Iv Characterization of L-Proline Sodium Sulphate Single Crystal 4.1 Structural Analysis (XRD)Document7 pagesChapter-Iv Characterization of L-Proline Sodium Sulphate Single Crystal 4.1 Structural Analysis (XRD)ramNo ratings yet
- TracStar 1800P7CF FADocument2 pagesTracStar 1800P7CF FAhennrynsNo ratings yet
- Chem 533 Lec03 PDFDocument42 pagesChem 533 Lec03 PDFJoseph MNo ratings yet
- Reflectivity Study of The 2-D Macroporous Silicon Photonic CrystalsDocument23 pagesReflectivity Study of The 2-D Macroporous Silicon Photonic CrystalsJosep Ferre-BorrullNo ratings yet
- E-Content Nonlinear Optical Applications of Phthalate CrystalsDocument54 pagesE-Content Nonlinear Optical Applications of Phthalate CrystalsbalaNo ratings yet
- An5426 Baseband CalculatorDocument13 pagesAn5426 Baseband CalculatorAdãodaLuzNo ratings yet
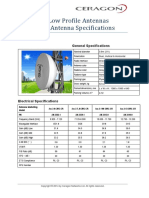
- 0.6m (2ft) Low Profile Antennas Microwave Antenna SpecificationsDocument15 pages0.6m (2ft) Low Profile Antennas Microwave Antenna SpecificationsDarwin Lopez AcevedoNo ratings yet
- OFS-100 FiberTrace Hilo 2Document2 pagesOFS-100 FiberTrace Hilo 2trubacoNo ratings yet
- CDS SlidesDocument9 pagesCDS SlidesswaroopNo ratings yet
- DF-P3HT SupportingDocument7 pagesDF-P3HT Supportingdkabra2ndNo ratings yet
- BH100 Bluray PresentationDocument80 pagesBH100 Bluray PresentationpasantitoNo ratings yet
- Lec 8 - Applications - CNTDocument76 pagesLec 8 - Applications - CNTMD SHAHIN ALOMNo ratings yet
- Department of Material Science Course: Material Study IDocument16 pagesDepartment of Material Science Course: Material Study IHelen LeNo ratings yet
- Enhancement of Sound Source Surrounded by Ambient Noise Using Pair of Microphone ArraysDocument4 pagesEnhancement of Sound Source Surrounded by Ambient Noise Using Pair of Microphone ArraysYusuke HiokaNo ratings yet
- Manual Espectrofotometro Parte 2Document30 pagesManual Espectrofotometro Parte 2Randy orhiel ramos alvarezNo ratings yet
- OFS-100 FiberTrace Hilo 1Document2 pagesOFS-100 FiberTrace Hilo 1trubacoNo ratings yet
- Cyp01x328yl Cyp01x32fxh 386Document1 pageCyp01x328yl Cyp01x32fxh 386ekawatNo ratings yet
- Otdr MakeDocument1 pageOtdr MakeekawatNo ratings yet
- Biomedical Optics IIDocument9 pagesBiomedical Optics IIAnonymous vcdqCTtS9No ratings yet
- Bti Passive ReferenceDocument1 pageBti Passive ReferenceAlberto CortezNo ratings yet
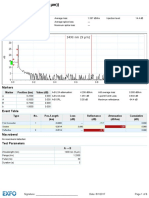
- Results: 0.0514 KM 0.080 DB 38.33 DBDocument6 pagesResults: 0.0514 KM 0.080 DB 38.33 DBAnonymous SefNnGNo ratings yet
- Lecture 3: Polymeric Biomaterials. Part 2 - Advanced Techniques For Analysis of MacromoleculesDocument26 pagesLecture 3: Polymeric Biomaterials. Part 2 - Advanced Techniques For Analysis of MacromoleculesNurul FarhahNo ratings yet
- SF-071803-C21 Data SheetDocument5 pagesSF-071803-C21 Data SheetAmir MeirNo ratings yet
- Box Properties: by Montani, Chocolates MontaniDocument6 pagesBox Properties: by Montani, Chocolates MontaniGilberto MontaniNo ratings yet
- Circular Dichroism LiteratureDocument39 pagesCircular Dichroism Literaturesuhas.lavhekarNo ratings yet
- Datasheet - Am-3-7 - 8-CIRC-CRDocument14 pagesDatasheet - Am-3-7 - 8-CIRC-CRkameshNo ratings yet
- Water Sensor From ChinaDocument60 pagesWater Sensor From ChinaAntoniusNo ratings yet
- Transmission Characteristics PDFDocument52 pagesTransmission Characteristics PDFDeepashreeKiran100% (1)
- New Methods For Nanoscale Fabrication and Characterization of Materials Primarily Using Electron and Ion BeamsDocument17 pagesNew Methods For Nanoscale Fabrication and Characterization of Materials Primarily Using Electron and Ion BeamsMaterials Research InstituteNo ratings yet
- Computed Radiographic Testing (CRT) Qualification Report: Rev. ADocument12 pagesComputed Radiographic Testing (CRT) Qualification Report: Rev. ATrung Tinh HoNo ratings yet
- Cuvette Guide 09-13Document27 pagesCuvette Guide 09-13Ramon Trinidad De la ONo ratings yet
- 1 4Document8 pages1 4AdheyfynmanNo ratings yet
- KW Oxides 2019-V9-Part-7Document28 pagesKW Oxides 2019-V9-Part-7qkzxbik734No ratings yet
- bc400 404 408 412 416 Data SheetDocument2 pagesbc400 404 408 412 416 Data SheetMario Javier GonzalesNo ratings yet
- Az p4620 PhotoresistDocument14 pagesAz p4620 Photoresist刘钢No ratings yet
- APE4517R0: Antenna SpecificationsDocument2 pagesAPE4517R0: Antenna SpecificationsКурбан УмархановNo ratings yet
- Hartman 1993 0248Document12 pagesHartman 1993 0248Particle Beam Physics LabNo ratings yet
- 158 MainDocument1 page158 MainekawatNo ratings yet
- Forensic DNA Fingerprinting: Using Restriction EnzymesDocument26 pagesForensic DNA Fingerprinting: Using Restriction EnzymesSyarifaha IhsanNo ratings yet
- Chrommatography Introduction-Handout-2023Document49 pagesChrommatography Introduction-Handout-2023Hien HoangNo ratings yet
- C. Lin, US-China 2009Document38 pagesC. Lin, US-China 2009Materials Research InstituteNo ratings yet
- TRS-398 JI Kielce 25-27 Nov 2004 PresentationDocument45 pagesTRS-398 JI Kielce 25-27 Nov 2004 PresentationGanesan RamanathanNo ratings yet
- Final A0 SMARANDACHE POSTER ECIS 2018Document1 pageFinal A0 SMARANDACHE POSTER ECIS 2018Adriana SmarandacheNo ratings yet
- GD Satcom 1184 C Ku Band VSATDocument1 pageGD Satcom 1184 C Ku Band VSATJawad MalikNo ratings yet
- 1000-036-Prodelin 1184 AntennaDocument1 page1000-036-Prodelin 1184 AntennaImran SadiqNo ratings yet
- Pa 09 02050Document7 pagesPa 09 02050Luciano PaoloNo ratings yet
- Refractive Index and Thickness Determinations Using A Dual-Path Mach-Zehnder Interferometer S.T. Lin, T.L. LinDocument5 pagesRefractive Index and Thickness Determinations Using A Dual-Path Mach-Zehnder Interferometer S.T. Lin, T.L. LinTinhNo ratings yet
- 372 mAINDocument1 page372 mAINekawatNo ratings yet
- Determination of Absorption SpectraDocument30 pagesDetermination of Absorption Spectrasay.elleseNo ratings yet
- IEE/CEEM Seminar October 21, 2009 Ucsb: Wesley Walker Toan Pho Dr. Rajeev Kumar Dr. Fulvio Brunetti Dr. Changduk YangDocument51 pagesIEE/CEEM Seminar October 21, 2009 Ucsb: Wesley Walker Toan Pho Dr. Rajeev Kumar Dr. Fulvio Brunetti Dr. Changduk YangMikaela MennenNo ratings yet
- Tutorial On ISO 10110 Optical Drawing StandardsDocument18 pagesTutorial On ISO 10110 Optical Drawing Standardstolga1234No ratings yet
- Atomic Force Microscopy For Materials First Edition PDFDocument35 pagesAtomic Force Microscopy For Materials First Edition PDFvivekdhand100% (1)
- Kinetika Pertumbuhan Mikroba Dalam Fermentasi PDFDocument40 pagesKinetika Pertumbuhan Mikroba Dalam Fermentasi PDFNano Music ProjectNo ratings yet
- A Fast and Efficient Approach To Prepare Starch Nanocrystals FromDocument7 pagesA Fast and Efficient Approach To Prepare Starch Nanocrystals FromLina Maria RodriguezNo ratings yet
- I UNIT Introduction of MicroscpeDocument11 pagesI UNIT Introduction of MicroscpeCokroNo ratings yet
- Test Bank For Microbiology The Human Experience First Edition First EditionDocument15 pagesTest Bank For Microbiology The Human Experience First Edition First Editionserenangahk5zNo ratings yet
- Optical Microscopy IntroductionDocument7 pagesOptical Microscopy IntroductionHuda AmirNo ratings yet
- Book of AbstractsDocument187 pagesBook of AbstractsVornicu Nicoleta100% (1)
- Corelative Fluorescence and Electron MicrosDocument10 pagesCorelative Fluorescence and Electron MicrosEmma StepinacNo ratings yet
- Light Microscopy: Basic Principles and Concepts: Carlo Bevilacqua Prevedel GroupDocument33 pagesLight Microscopy: Basic Principles and Concepts: Carlo Bevilacqua Prevedel Groupalvise30No ratings yet
- E-Conservation Magazine - 10Document82 pagesE-Conservation Magazine - 10conservatorsNo ratings yet
- Practical NotebookDocument20 pagesPractical NotebookhemalerashidNo ratings yet
- Cell Structure and FunctionDocument4 pagesCell Structure and FunctionRidham JainNo ratings yet
- B Acte CheckDocument126 pagesB Acte CheckMarie LlanesNo ratings yet
- Nanocharacterisation, 2007, p.319 PDFDocument319 pagesNanocharacterisation, 2007, p.319 PDFMaria Ana Vulcu100% (1)
- DOMINGO, Catherine M.-Laboratory Activity - 2 - Compound MicroscopeDocument4 pagesDOMINGO, Catherine M.-Laboratory Activity - 2 - Compound MicroscopeJohn Carl DomingoNo ratings yet
- Parts of The CellDocument26 pagesParts of The CellMarion AlbertoNo ratings yet
- Advanced Microscopy Simplifi Ed: Bx53M/BxfmDocument28 pagesAdvanced Microscopy Simplifi Ed: Bx53M/BxfmRepresentaciones y Distribuciones FALNo ratings yet
- Cells: Subject: Integrated Science Level: Second Form Duration: 3-5 Weeks January 21st - February 22, 2019Document30 pagesCells: Subject: Integrated Science Level: Second Form Duration: 3-5 Weeks January 21st - February 22, 2019Alberto SholNo ratings yet
- What Is BacteriaDocument33 pagesWhat Is Bacteriaapi-19862227100% (1)
- Tortora Micro Chapter 13 Flashcards - QuizletDocument10 pagesTortora Micro Chapter 13 Flashcards - QuizletdalgomNo ratings yet
- Candida Sinteza ImaginiDocument30 pagesCandida Sinteza ImaginiSandra PopescuNo ratings yet
- Prokaryotic and Eukaryotic CellDocument19 pagesProkaryotic and Eukaryotic CellReanne MeregildoNo ratings yet
- Course OutlineDocument19 pagesCourse OutlineHernando VaydalNo ratings yet
- Principles of Antimicrobial ChemotherapyDocument3 pagesPrinciples of Antimicrobial ChemotherapySteven miles PunzalanNo ratings yet
- Nitrobacter WinogradskyDocument7 pagesNitrobacter WinogradskyFerryKurniawanNo ratings yet
- FemtoFiber Ultra 920 TOPTICA - Released2Document4 pagesFemtoFiber Ultra 920 TOPTICA - Released2Green LemonNo ratings yet






















































































Download as pdf or txt
You might also like
- Biology 2nd Lab Report (Onion Cell)Document4 pagesBiology 2nd Lab Report (Onion Cell)Johanna Haludilu89% (18)
- Cells Exploration ActivitiesDocument32 pagesCells Exploration ActivitiesSay KhoNo ratings yet
- Cobas U 701 Operators Manual EN (Initial Version) 1.0.0 ENDocument238 pagesCobas U 701 Operators Manual EN (Initial Version) 1.0.0 ENRicky100% (1)
- Ja4c05178 Si 001Document9 pagesJa4c05178 Si 001Yong-Sheng WeiNo ratings yet
- La0c02115 Si 001Document13 pagesLa0c02115 Si 001peeyush kumarNo ratings yet
- Basis of Computed Radiography & PACS: Digital Film-ScreenDocument28 pagesBasis of Computed Radiography & PACS: Digital Film-ScreenAmar MohammedNo ratings yet
- Dyes SupportDocument7 pagesDyes SupportawaisNo ratings yet
- NTN Pauhlimo-Ombilin Core12Document2 pagesNTN Pauhlimo-Ombilin Core12rahman.ab1212No ratings yet
- Silva 2014CO2解离的微波脉冲放电的光学表征Document14 pagesSilva 2014CO2解离的微波脉冲放电的光学表征yuan MoNo ratings yet
- ImagingDocument29 pagesImagingDyhoiNo ratings yet
- 12mihailidis ElectronBeamDosimetryDocument92 pages12mihailidis ElectronBeamDosimetryitumNo ratings yet
- Tribological Properties of Diamond-Like Carbon Films Produced by Different Deposition TechniquesDocument5 pagesTribological Properties of Diamond-Like Carbon Films Produced by Different Deposition TechniquesEngels OchoaNo ratings yet
- Chapter-Iv Characterization of L-Proline Sodium Sulphate Single Crystal 4.1 Structural Analysis (XRD)Document7 pagesChapter-Iv Characterization of L-Proline Sodium Sulphate Single Crystal 4.1 Structural Analysis (XRD)ramNo ratings yet
- TracStar 1800P7CF FADocument2 pagesTracStar 1800P7CF FAhennrynsNo ratings yet
- Chem 533 Lec03 PDFDocument42 pagesChem 533 Lec03 PDFJoseph MNo ratings yet
- Reflectivity Study of The 2-D Macroporous Silicon Photonic CrystalsDocument23 pagesReflectivity Study of The 2-D Macroporous Silicon Photonic CrystalsJosep Ferre-BorrullNo ratings yet
- E-Content Nonlinear Optical Applications of Phthalate CrystalsDocument54 pagesE-Content Nonlinear Optical Applications of Phthalate CrystalsbalaNo ratings yet
- An5426 Baseband CalculatorDocument13 pagesAn5426 Baseband CalculatorAdãodaLuzNo ratings yet
- 0.6m (2ft) Low Profile Antennas Microwave Antenna SpecificationsDocument15 pages0.6m (2ft) Low Profile Antennas Microwave Antenna SpecificationsDarwin Lopez AcevedoNo ratings yet
- OFS-100 FiberTrace Hilo 2Document2 pagesOFS-100 FiberTrace Hilo 2trubacoNo ratings yet
- CDS SlidesDocument9 pagesCDS SlidesswaroopNo ratings yet
- DF-P3HT SupportingDocument7 pagesDF-P3HT Supportingdkabra2ndNo ratings yet
- BH100 Bluray PresentationDocument80 pagesBH100 Bluray PresentationpasantitoNo ratings yet
- Lec 8 - Applications - CNTDocument76 pagesLec 8 - Applications - CNTMD SHAHIN ALOMNo ratings yet
- Department of Material Science Course: Material Study IDocument16 pagesDepartment of Material Science Course: Material Study IHelen LeNo ratings yet
- Enhancement of Sound Source Surrounded by Ambient Noise Using Pair of Microphone ArraysDocument4 pagesEnhancement of Sound Source Surrounded by Ambient Noise Using Pair of Microphone ArraysYusuke HiokaNo ratings yet
- Manual Espectrofotometro Parte 2Document30 pagesManual Espectrofotometro Parte 2Randy orhiel ramos alvarezNo ratings yet
- OFS-100 FiberTrace Hilo 1Document2 pagesOFS-100 FiberTrace Hilo 1trubacoNo ratings yet
- Cyp01x328yl Cyp01x32fxh 386Document1 pageCyp01x328yl Cyp01x32fxh 386ekawatNo ratings yet
- Otdr MakeDocument1 pageOtdr MakeekawatNo ratings yet
- Biomedical Optics IIDocument9 pagesBiomedical Optics IIAnonymous vcdqCTtS9No ratings yet
- Bti Passive ReferenceDocument1 pageBti Passive ReferenceAlberto CortezNo ratings yet
- Results: 0.0514 KM 0.080 DB 38.33 DBDocument6 pagesResults: 0.0514 KM 0.080 DB 38.33 DBAnonymous SefNnGNo ratings yet
- Lecture 3: Polymeric Biomaterials. Part 2 - Advanced Techniques For Analysis of MacromoleculesDocument26 pagesLecture 3: Polymeric Biomaterials. Part 2 - Advanced Techniques For Analysis of MacromoleculesNurul FarhahNo ratings yet
- SF-071803-C21 Data SheetDocument5 pagesSF-071803-C21 Data SheetAmir MeirNo ratings yet
- Box Properties: by Montani, Chocolates MontaniDocument6 pagesBox Properties: by Montani, Chocolates MontaniGilberto MontaniNo ratings yet
- Circular Dichroism LiteratureDocument39 pagesCircular Dichroism Literaturesuhas.lavhekarNo ratings yet
- Datasheet - Am-3-7 - 8-CIRC-CRDocument14 pagesDatasheet - Am-3-7 - 8-CIRC-CRkameshNo ratings yet
- Water Sensor From ChinaDocument60 pagesWater Sensor From ChinaAntoniusNo ratings yet
- Transmission Characteristics PDFDocument52 pagesTransmission Characteristics PDFDeepashreeKiran100% (1)
- New Methods For Nanoscale Fabrication and Characterization of Materials Primarily Using Electron and Ion BeamsDocument17 pagesNew Methods For Nanoscale Fabrication and Characterization of Materials Primarily Using Electron and Ion BeamsMaterials Research InstituteNo ratings yet
- Computed Radiographic Testing (CRT) Qualification Report: Rev. ADocument12 pagesComputed Radiographic Testing (CRT) Qualification Report: Rev. ATrung Tinh HoNo ratings yet
- Cuvette Guide 09-13Document27 pagesCuvette Guide 09-13Ramon Trinidad De la ONo ratings yet
- 1 4Document8 pages1 4AdheyfynmanNo ratings yet
- KW Oxides 2019-V9-Part-7Document28 pagesKW Oxides 2019-V9-Part-7qkzxbik734No ratings yet
- bc400 404 408 412 416 Data SheetDocument2 pagesbc400 404 408 412 416 Data SheetMario Javier GonzalesNo ratings yet
- Az p4620 PhotoresistDocument14 pagesAz p4620 Photoresist刘钢No ratings yet
- APE4517R0: Antenna SpecificationsDocument2 pagesAPE4517R0: Antenna SpecificationsКурбан УмархановNo ratings yet
- Hartman 1993 0248Document12 pagesHartman 1993 0248Particle Beam Physics LabNo ratings yet
- 158 MainDocument1 page158 MainekawatNo ratings yet
- Forensic DNA Fingerprinting: Using Restriction EnzymesDocument26 pagesForensic DNA Fingerprinting: Using Restriction EnzymesSyarifaha IhsanNo ratings yet
- Chrommatography Introduction-Handout-2023Document49 pagesChrommatography Introduction-Handout-2023Hien HoangNo ratings yet
- C. Lin, US-China 2009Document38 pagesC. Lin, US-China 2009Materials Research InstituteNo ratings yet
- TRS-398 JI Kielce 25-27 Nov 2004 PresentationDocument45 pagesTRS-398 JI Kielce 25-27 Nov 2004 PresentationGanesan RamanathanNo ratings yet
- Final A0 SMARANDACHE POSTER ECIS 2018Document1 pageFinal A0 SMARANDACHE POSTER ECIS 2018Adriana SmarandacheNo ratings yet
- GD Satcom 1184 C Ku Band VSATDocument1 pageGD Satcom 1184 C Ku Band VSATJawad MalikNo ratings yet
- 1000-036-Prodelin 1184 AntennaDocument1 page1000-036-Prodelin 1184 AntennaImran SadiqNo ratings yet
- Pa 09 02050Document7 pagesPa 09 02050Luciano PaoloNo ratings yet
- Refractive Index and Thickness Determinations Using A Dual-Path Mach-Zehnder Interferometer S.T. Lin, T.L. LinDocument5 pagesRefractive Index and Thickness Determinations Using A Dual-Path Mach-Zehnder Interferometer S.T. Lin, T.L. LinTinhNo ratings yet
- 372 mAINDocument1 page372 mAINekawatNo ratings yet
- Determination of Absorption SpectraDocument30 pagesDetermination of Absorption Spectrasay.elleseNo ratings yet
- IEE/CEEM Seminar October 21, 2009 Ucsb: Wesley Walker Toan Pho Dr. Rajeev Kumar Dr. Fulvio Brunetti Dr. Changduk YangDocument51 pagesIEE/CEEM Seminar October 21, 2009 Ucsb: Wesley Walker Toan Pho Dr. Rajeev Kumar Dr. Fulvio Brunetti Dr. Changduk YangMikaela MennenNo ratings yet
- Tutorial On ISO 10110 Optical Drawing StandardsDocument18 pagesTutorial On ISO 10110 Optical Drawing Standardstolga1234No ratings yet
- Atomic Force Microscopy For Materials First Edition PDFDocument35 pagesAtomic Force Microscopy For Materials First Edition PDFvivekdhand100% (1)
- Kinetika Pertumbuhan Mikroba Dalam Fermentasi PDFDocument40 pagesKinetika Pertumbuhan Mikroba Dalam Fermentasi PDFNano Music ProjectNo ratings yet
- A Fast and Efficient Approach To Prepare Starch Nanocrystals FromDocument7 pagesA Fast and Efficient Approach To Prepare Starch Nanocrystals FromLina Maria RodriguezNo ratings yet
- I UNIT Introduction of MicroscpeDocument11 pagesI UNIT Introduction of MicroscpeCokroNo ratings yet
- Test Bank For Microbiology The Human Experience First Edition First EditionDocument15 pagesTest Bank For Microbiology The Human Experience First Edition First Editionserenangahk5zNo ratings yet
- Optical Microscopy IntroductionDocument7 pagesOptical Microscopy IntroductionHuda AmirNo ratings yet
- Book of AbstractsDocument187 pagesBook of AbstractsVornicu Nicoleta100% (1)
- Corelative Fluorescence and Electron MicrosDocument10 pagesCorelative Fluorescence and Electron MicrosEmma StepinacNo ratings yet
- Light Microscopy: Basic Principles and Concepts: Carlo Bevilacqua Prevedel GroupDocument33 pagesLight Microscopy: Basic Principles and Concepts: Carlo Bevilacqua Prevedel Groupalvise30No ratings yet
- E-Conservation Magazine - 10Document82 pagesE-Conservation Magazine - 10conservatorsNo ratings yet
- Practical NotebookDocument20 pagesPractical NotebookhemalerashidNo ratings yet
- Cell Structure and FunctionDocument4 pagesCell Structure and FunctionRidham JainNo ratings yet
- B Acte CheckDocument126 pagesB Acte CheckMarie LlanesNo ratings yet
- Nanocharacterisation, 2007, p.319 PDFDocument319 pagesNanocharacterisation, 2007, p.319 PDFMaria Ana Vulcu100% (1)
- DOMINGO, Catherine M.-Laboratory Activity - 2 - Compound MicroscopeDocument4 pagesDOMINGO, Catherine M.-Laboratory Activity - 2 - Compound MicroscopeJohn Carl DomingoNo ratings yet
- Parts of The CellDocument26 pagesParts of The CellMarion AlbertoNo ratings yet
- Advanced Microscopy Simplifi Ed: Bx53M/BxfmDocument28 pagesAdvanced Microscopy Simplifi Ed: Bx53M/BxfmRepresentaciones y Distribuciones FALNo ratings yet
- Cells: Subject: Integrated Science Level: Second Form Duration: 3-5 Weeks January 21st - February 22, 2019Document30 pagesCells: Subject: Integrated Science Level: Second Form Duration: 3-5 Weeks January 21st - February 22, 2019Alberto SholNo ratings yet
- What Is BacteriaDocument33 pagesWhat Is Bacteriaapi-19862227100% (1)
- Tortora Micro Chapter 13 Flashcards - QuizletDocument10 pagesTortora Micro Chapter 13 Flashcards - QuizletdalgomNo ratings yet
- Candida Sinteza ImaginiDocument30 pagesCandida Sinteza ImaginiSandra PopescuNo ratings yet
- Prokaryotic and Eukaryotic CellDocument19 pagesProkaryotic and Eukaryotic CellReanne MeregildoNo ratings yet
- Course OutlineDocument19 pagesCourse OutlineHernando VaydalNo ratings yet
- Principles of Antimicrobial ChemotherapyDocument3 pagesPrinciples of Antimicrobial ChemotherapySteven miles PunzalanNo ratings yet
- Nitrobacter WinogradskyDocument7 pagesNitrobacter WinogradskyFerryKurniawanNo ratings yet
- FemtoFiber Ultra 920 TOPTICA - Released2Document4 pagesFemtoFiber Ultra 920 TOPTICA - Released2Green LemonNo ratings yet