Download as pdf or txt
You might also like
- Radio Ti Kan - Audi Concert II RadioDocument2 pagesRadio Ti Kan - Audi Concert II Radiogeorge_adi_2200No ratings yet
- Dork Diaries 14 Tales From A NOT SO Best Friend Sneak Peek PDFDocument19 pagesDork Diaries 14 Tales From A NOT SO Best Friend Sneak Peek PDFTrisha Rai33% (9)
- BrainVision PyCorder UM PDFDocument86 pagesBrainVision PyCorder UM PDFrm7029No ratings yet
- Thermal Stability of I.T. at SiteDocument4 pagesThermal Stability of I.T. at SiteBalubhai KarsanbhaiNo ratings yet
- Vidyanidhi Info Tech Academy: PG DAC Question BankDocument23 pagesVidyanidhi Info Tech Academy: PG DAC Question BankVishal YadavNo ratings yet
- Kapitein 2005Document5 pagesKapitein 2005liliana-contrerasNo ratings yet
- Neutralizing Antibodies in Patients With Severe Acute Respiratory Syndrome-Associated Coronavirus InfectionDocument8 pagesNeutralizing Antibodies in Patients With Severe Acute Respiratory Syndrome-Associated Coronavirus InfectionNicolas CasanovaNo ratings yet
- Route of Vaccine Administration Alters Antigen Trafficking But Not Innate or Adaptive ImmunityDocument16 pagesRoute of Vaccine Administration Alters Antigen Trafficking But Not Innate or Adaptive Immunityemmanuel AndemNo ratings yet
- DNA Vaccines: Ready For Prime Time?: Michele A. Kutzler and David B. WeinerDocument13 pagesDNA Vaccines: Ready For Prime Time?: Michele A. Kutzler and David B. WeinerantNo ratings yet
- Alpha-C-galactosylceramide As An Adjuvant For A Live Attenuated in Uenza Virus VaccineDocument9 pagesAlpha-C-galactosylceramide As An Adjuvant For A Live Attenuated in Uenza Virus VaccineLeandro CarreñoNo ratings yet
- Grammatical CohesionDocument6 pagesGrammatical CohesionMacarena Diaz CasadoNo ratings yet
- Science Abc6284 FullDocument11 pagesScience Abc6284 FullPatrick CoNo ratings yet
- Review 2010Document2 pagesReview 2010Igor VainerNo ratings yet
- Ijms 21 02903Document16 pagesIjms 21 02903vuongquynh290900No ratings yet
- Sars-Cov-2 Mrna Vaccines Induce Persistent Human Germinal Centre ResponsesDocument22 pagesSars-Cov-2 Mrna Vaccines Induce Persistent Human Germinal Centre ResponsesPedro Andrés R. SánchezNo ratings yet
- COVID-19 Makes B Cells Forget, But T Cells Remember: PreviewsDocument3 pagesCOVID-19 Makes B Cells Forget, But T Cells Remember: PreviewsRamy Refaat LewisNo ratings yet
- Main PDFDocument12 pagesMain PDFLuz Betty Alejo MamaniNo ratings yet
- Gut Microbiome and anti-PD1Document9 pagesGut Microbiome and anti-PD1Suad AjabNo ratings yet
- Surrogates For The Study of Norovirus Stability AnDocument6 pagesSurrogates For The Study of Norovirus Stability Andanasli7607245No ratings yet
- SARS-CoV-2 Paper-ScienceDocument12 pagesSARS-CoV-2 Paper-ScienceCavin PengNo ratings yet
- Articles: BackgroundDocument12 pagesArticles: BackgroundBudi Haryadi PNo ratings yet
- Increased Humoral Immunity by DNA Vaccination Using An A-Tocopherol-Based AdjuvantDocument8 pagesIncreased Humoral Immunity by DNA Vaccination Using An A-Tocopherol-Based AdjuvantFakultas Kedokteran UnhanNo ratings yet
- Chlorovirus ATCV-1 Is Part of The Human Oropharyngeal Virome and Is Associated With Changes in Cognitive Functions in Humans and MiceDocument6 pagesChlorovirus ATCV-1 Is Part of The Human Oropharyngeal Virome and Is Associated With Changes in Cognitive Functions in Humans and Micebwchamp1No ratings yet
- 1996 Mesri JemDocument6 pages1996 Mesri JemleandrosarvanitakisNo ratings yet
- Inducible Rubicon Facilitates Viral Replication by Antagonizing Interferon ProductionDocument14 pagesInducible Rubicon Facilitates Viral Replication by Antagonizing Interferon ProductionfreddyNo ratings yet
- Studies in Humanized Mice and Convalescent Humans Yield A Sars-Cov-2 Antibody CocktailDocument7 pagesStudies in Humanized Mice and Convalescent Humans Yield A Sars-Cov-2 Antibody CocktailscsldvNo ratings yet
- 1 s2.0 S0166354209002162 MainDocument5 pages1 s2.0 S0166354209002162 MainTope BondocNo ratings yet
- Zhao Et Al 2013Document10 pagesZhao Et Al 2013lionyginting369No ratings yet
- 1 - Hudson 2003. Engineered Antibodies. EditadoDocument6 pages1 - Hudson 2003. Engineered Antibodies. EditadoIara RothNo ratings yet
- Journal Neuro 6Document22 pagesJournal Neuro 6Najla ShuaibNo ratings yet
- The SARS-Coronavirus-Host Interactome: Identification of Cyclophilins As Target For Pan-Coronavirus InhibitorsDocument15 pagesThe SARS-Coronavirus-Host Interactome: Identification of Cyclophilins As Target For Pan-Coronavirus Inhibitorsfaris daboolNo ratings yet
- 440 FullDocument3 pages440 Fullantarex06No ratings yet
- 1 s2.0 S1569199321019652 MainDocument1 page1 s2.0 S1569199321019652 MainAnita DewiNo ratings yet
- Seminario 2-2022-1Document3 pagesSeminario 2-2022-1Elias Gerardo Pardo LlamoccaNo ratings yet
- 17 Rapid, Robust, and Sustainable Antibody Responses To mRNA COVID-19 Vaccine in Convalescent COVID-19 IndividualsDocument14 pages17 Rapid, Robust, and Sustainable Antibody Responses To mRNA COVID-19 Vaccine in Convalescent COVID-19 Individualsnadhia rahdaNo ratings yet
- Construction and Evaluation of A Novel Bacillus Subtilis Spores-Based Enterovirus 71 VaccineDocument9 pagesConstruction and Evaluation of A Novel Bacillus Subtilis Spores-Based Enterovirus 71 VaccineKhoi Nguyen Nguyen HoangNo ratings yet
- Antibody Landscapes After Influenza Virus Infection or VaccinationDocument6 pagesAntibody Landscapes After Influenza Virus Infection or VaccinationManuel Parra CardeñaNo ratings yet
- Test 11Document14 pagesTest 11CiprianNo ratings yet
- WissenshaftDocument11 pagesWissenshaftkacagj bajazzoNo ratings yet
- 1 s2.0 S1074761320303265 MainDocument17 pages1 s2.0 S1074761320303265 MainAleksandar DimkovskiNo ratings yet
- MMC 5Document21 pagesMMC 5Andreea AlexandruNo ratings yet
- A Sars Like Cluster of Circulating Bat Coronaviruses Shows Potential For Human EmergenceDocument17 pagesA Sars Like Cluster of Circulating Bat Coronaviruses Shows Potential For Human EmergencedavinmaximusNo ratings yet
- 1 s2.0 S1569199321019548 MainDocument1 page1 s2.0 S1569199321019548 MainAnita DewiNo ratings yet
- Gilbert2005 Studi HivDocument12 pagesGilbert2005 Studi HivErnawati HidayatNo ratings yet
- Literature Review 3.1 Dengue VaccineDocument5 pagesLiterature Review 3.1 Dengue VaccineabdulNo ratings yet
- 10 1126@science Aau6481Document5 pages10 1126@science Aau6481nnbfgwnrhbNo ratings yet
- Epigenetic Silencing of SETDB1Document28 pagesEpigenetic Silencing of SETDB1Dario AnobileNo ratings yet
- Cov Immunotherapy 2Document2 pagesCov Immunotherapy 2lakshminivas PingaliNo ratings yet
- 1 s2.0 S0264410X20312597 MainDocument10 pages1 s2.0 S0264410X20312597 MainLixia WeiNo ratings yet
- An Oral SARS-CoV-2 Mpro Inhibitor Clinical Candidate For The Treatment of COVID-19Document8 pagesAn Oral SARS-CoV-2 Mpro Inhibitor Clinical Candidate For The Treatment of COVID-19Leonel RiveraNo ratings yet
- Enterobacter Cloacae: Carbapenem-Resistant in Patients From The US Veterans Health Administration, 2006-2015Document3 pagesEnterobacter Cloacae: Carbapenem-Resistant in Patients From The US Veterans Health Administration, 2006-2015Mayang WulanNo ratings yet
- ICP34.5 Deleted Herpes Simplex Virus With Enhanced Oncolytic, Immune Stimulating, and Anti-Tumour PropertiesDocument12 pagesICP34.5 Deleted Herpes Simplex Virus With Enhanced Oncolytic, Immune Stimulating, and Anti-Tumour PropertiesmostafaNo ratings yet
- 140794.3 20210324105625 CoveredDocument16 pages140794.3 20210324105625 CoveredBruno RamírezNo ratings yet
- Heterogenous Humoral and Cellular Immune Responses With Distinct Trajectories postSARS-CoV-2 Infection in A Population-Based CohortDocument16 pagesHeterogenous Humoral and Cellular Immune Responses With Distinct Trajectories postSARS-CoV-2 Infection in A Population-Based CohortYuzhuo WangNo ratings yet
- MOT 2020 - Transcriptional Profiles of Adjuvanted Hepatitis B Vaccines Display Variable Interindividual Homogeneity But A Shared Core SignatureDocument14 pagesMOT 2020 - Transcriptional Profiles of Adjuvanted Hepatitis B Vaccines Display Variable Interindividual Homogeneity But A Shared Core SignatureJoão Pedro NunesNo ratings yet
- Dynamics of Bovine Spleen Cell Populations DuringDocument12 pagesDynamics of Bovine Spleen Cell Populations DuringMony ElbazNo ratings yet
- Microvesicles Are A Source of Contaminating Cellular Proteins Found in Purified HIV-1 PreparationsDocument11 pagesMicrovesicles Are A Source of Contaminating Cellular Proteins Found in Purified HIV-1 PreparationsPipirrin GonzalesNo ratings yet
- A Cultured Affair HSV Latency and Reactivation in Neurons Jan2014Document8 pagesA Cultured Affair HSV Latency and Reactivation in Neurons Jan2014Ana-Maria DavidNo ratings yet
- Identification of Cathepsin B in The Razor Clam Sinonovacula Constricta and Its Role in Innate Immune ResponsesDocument6 pagesIdentification of Cathepsin B in The Razor Clam Sinonovacula Constricta and Its Role in Innate Immune ResponsesRizka Rahmana PutriNo ratings yet
- Articulo Científico para LeerDocument27 pagesArticulo Científico para LeerwilverNo ratings yet
- Historical Overview of The Cephalosporin Spectrum: Four Generations of Structural EvolutionDocument8 pagesHistorical Overview of The Cephalosporin Spectrum: Four Generations of Structural EvolutionBagus FitraNo ratings yet
- Jama Abbasi 2021 MN 210043 1627852635.57854Document2 pagesJama Abbasi 2021 MN 210043 1627852635.57854anggaririnNo ratings yet
- Mucoadhesive Glycol Chitosan Nanoparticles For Intranasal Delivery of Hepatitis B Vaccine Enhancement of Mucosal and Systemic Immune ResponseDocument11 pagesMucoadhesive Glycol Chitosan Nanoparticles For Intranasal Delivery of Hepatitis B Vaccine Enhancement of Mucosal and Systemic Immune ResponseKhalifatul Aulia AsyuriyahNo ratings yet
- COVID-19 Vaccine Design: The Janus Face of Immune EnhancementDocument2 pagesCOVID-19 Vaccine Design: The Janus Face of Immune EnhancementPratiwi PuspanegaraNo ratings yet
- Molecular Basis of Biological Degradative processesFrom EverandMolecular Basis of Biological Degradative processesRichard BerlinNo ratings yet
- 10 - Bronchiolitis Clinics and Medical TreatmentDocument12 pages10 - Bronchiolitis Clinics and Medical TreatmentamyNo ratings yet
- Seroprevalence of anti-SARS-CoV-2 Antibodies in COVID-19 Patients and Healthy Volunteers Up To 6 Months Post Disease OnsetDocument16 pagesSeroprevalence of anti-SARS-CoV-2 Antibodies in COVID-19 Patients and Healthy Volunteers Up To 6 Months Post Disease OnsetamyNo ratings yet
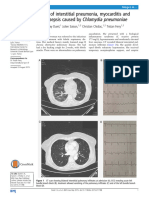
- A Case of Interstitial Pneumonia, Myocarditis Caused by C.pneumoniaeDocument2 pagesA Case of Interstitial Pneumonia, Myocarditis Caused by C.pneumoniaeamyNo ratings yet
- Vaccination Contre La Coqueluche Au Sein de Trois Populations D'adultes France2017Document7 pagesVaccination Contre La Coqueluche Au Sein de Trois Populations D'adultes France2017amyNo ratings yet
- Risk Factors For Community-Acquired Pneumonia in Adults, A Systematic Review of Observational Studies 2017Document13 pagesRisk Factors For Community-Acquired Pneumonia in Adults, A Systematic Review of Observational Studies 2017amyNo ratings yet
- Advances in Community-Acquired Pneumonia 2020Document11 pagesAdvances in Community-Acquired Pneumonia 2020amyNo ratings yet
- List of Shortlisted Candidates For InterviewDocument19 pagesList of Shortlisted Candidates For InterviewJenitaNo ratings yet
- Preparing For GO!Enterprise MDM On-Demand Service: This Guide Provides Information On - .Document11 pagesPreparing For GO!Enterprise MDM On-Demand Service: This Guide Provides Information On - .Jarbas SpNo ratings yet
- Smartphone Apps in Education: Students Create Videos To Teach Smartphone Use As Tool For LearningDocument11 pagesSmartphone Apps in Education: Students Create Videos To Teach Smartphone Use As Tool For LearningKossuthos BélesNo ratings yet
- Table Manners in A Formal Restaurant. 1.: Make Good Use of Your Napkin. Place Your Napkin in Your Lap Immediately UponDocument2 pagesTable Manners in A Formal Restaurant. 1.: Make Good Use of Your Napkin. Place Your Napkin in Your Lap Immediately UponAllen KateNo ratings yet
- R.A 10171Document2 pagesR.A 10171Hazel Anne MarianoNo ratings yet
- Appendix A: A.1. Three Phase SeparatorDocument48 pagesAppendix A: A.1. Three Phase SeparatorNhaaaeyNo ratings yet
- Globo PLC H1 2015 ReportDocument20 pagesGlobo PLC H1 2015 Reportjenkins-sacadonaNo ratings yet
- 7plus Unit 1 Lesson 1 Opposite Quantities Combine To Make ZeroDocument6 pages7plus Unit 1 Lesson 1 Opposite Quantities Combine To Make Zeroapi-261894355No ratings yet
- 5.3.1.2 Packet Tracer - Skills Integration Challenge InstructionsDocument2 pages5.3.1.2 Packet Tracer - Skills Integration Challenge InstructionsTom0% (1)
- Drones 06 00019 v2Document19 pagesDrones 06 00019 v2GERALD BRANDI CAINICELA AQUINONo ratings yet
- Graphic Communication TechniquesDocument41 pagesGraphic Communication TechniquesKim TruongNo ratings yet
- Solid Waste Management Dissartation Report (Jun 2013)Document60 pagesSolid Waste Management Dissartation Report (Jun 2013)Anonymous eTFc35tQ2No ratings yet
- Assignment CHPT 12Document3 pagesAssignment CHPT 12Sultan LimitNo ratings yet
- Teaching With Simulations: What Are Instructional Simulations?Document4 pagesTeaching With Simulations: What Are Instructional Simulations?Norazmah BachokNo ratings yet
- Port of Newcastle - Hydrogen PlansDocument6 pagesPort of Newcastle - Hydrogen PlansMatthew KellyNo ratings yet
- Research Seminar Course Outline GlobalmarketingDocument3 pagesResearch Seminar Course Outline GlobalmarketingSaid Ahmed100% (1)
- Reliable Distance Protection: Get Precise Line Parameters With The CPC 100 + CP CU1Document3 pagesReliable Distance Protection: Get Precise Line Parameters With The CPC 100 + CP CU1Jonas PedrosoNo ratings yet
- Unit 1 Adr WordDocument4 pagesUnit 1 Adr WordmilkymilkyNo ratings yet
- Youtubedownloader CodeDocument3 pagesYoutubedownloader CodeYusa 85No ratings yet
- Introduction To Instrumentation and Control EngineeringDocument38 pagesIntroduction To Instrumentation and Control EngineeringJames GupoNo ratings yet
- Combination StrategyDocument5 pagesCombination StrategyNadya ShaminiNo ratings yet
- Advantages and Disadvantages of Integrated CircuitDocument5 pagesAdvantages and Disadvantages of Integrated CircuitBappy HossainNo ratings yet
- A Study of Big Data Characteristics: October 2016Document5 pagesA Study of Big Data Characteristics: October 2016Johnny AndreatyNo ratings yet
- Tenders Soon For 3 Hotels Near Navi Mumbai Airport - Times of IndiaDocument2 pagesTenders Soon For 3 Hotels Near Navi Mumbai Airport - Times of IndiaRasika WankhadeNo ratings yet