
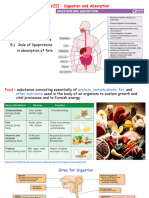
Metabolism
Metabolism
You might also like
- Biochemistry II End of Semester Exam QuestionsDocument6 pagesBiochemistry II End of Semester Exam Questionsnima00No ratings yet
- Biochemistry MCQsDocument19 pagesBiochemistry MCQsschxzerrydawn100% (1)
- Testbank 4e CH14Document19 pagesTestbank 4e CH14somasudeshna100% (5)
- UntitledDocument14 pagesUntitledSyafrianda PanggabeanNo ratings yet
- Unit 8 Digestion and AbsorptionDocument33 pagesUnit 8 Digestion and AbsorptionGlory MuedaNo ratings yet
- Biology M5 Cellular RespirationDocument30 pagesBiology M5 Cellular RespirationpatrickkayeNo ratings yet
- Docs 2Document13 pagesDocs 2Mary Rose CuentasNo ratings yet
- 04 Energy TransformationDocument53 pages04 Energy TransformationlyleandreihendersonNo ratings yet
- Lecture 02. Common Metabolic Pathways. Biological Oxidation. Tissue Respiration. Oxidative PhosphorylationDocument53 pagesLecture 02. Common Metabolic Pathways. Biological Oxidation. Tissue Respiration. Oxidative PhosphorylationВіталій Михайлович НечипорукNo ratings yet
- MODULE - FINALS - Biochem - 1Document5 pagesMODULE - FINALS - Biochem - 1Cyril CauilanNo ratings yet
- GEN BIO RevsDocument7 pagesGEN BIO RevsacglianeNo ratings yet
- Pre Test: Prepared By: Patricia PradoDocument51 pagesPre Test: Prepared By: Patricia PradoLovely AlosadoNo ratings yet
- Module 12 PDFDocument13 pagesModule 12 PDFJohn Bernard VeloriaNo ratings yet
- Biochemistry (Metabolism) by Moses KDocument405 pagesBiochemistry (Metabolism) by Moses Khaamajansi86% (7)
- Module 8 Metabolic Pathways Final 1Document56 pagesModule 8 Metabolic Pathways Final 1Susana N peNo ratings yet
- 3 CarbohydratesDocument12 pages3 CarbohydratesconcepcionsofiakarylleNo ratings yet
- How Animals Survive Lesson 5Document30 pagesHow Animals Survive Lesson 5Ellah LaguismaNo ratings yet
- How Cells Release Energy Part 1Document14 pagesHow Cells Release Energy Part 1Jiro SarmientoNo ratings yet
- 05 F4 Biology Chap 6Document19 pages05 F4 Biology Chap 6Benjamin TeeNo ratings yet
- The Biochemistry of Digestion, Absorption and Detoxification by Prof. Dr. Hedef D. El-YassinDocument64 pagesThe Biochemistry of Digestion, Absorption and Detoxification by Prof. Dr. Hedef D. El-YassinSayhidoen CepexNo ratings yet
- Agri-501 Biochemistry 2021-2022: Doc. Regina P. ClavelDocument36 pagesAgri-501 Biochemistry 2021-2022: Doc. Regina P. ClavelIvan Jhon AnamNo ratings yet
- Biology The Essentials 1st Edition Hoefnagels Solutions Manual DownloadDocument15 pagesBiology The Essentials 1st Edition Hoefnagels Solutions Manual DownloadGraham Hissem100% (26)
- Carbohydrates Part 2 - MetabolismsDocument60 pagesCarbohydrates Part 2 - MetabolismsNikka Mia AbadiesNo ratings yet
- Metabolic ProcessesDocument6 pagesMetabolic ProcessesMark Andrian PontanalNo ratings yet
- Last Cellmolecule ReportDocument10 pagesLast Cellmolecule Reportapi-638362671No ratings yet
- Life Science Module 4 Lesson 5 and 6Document38 pagesLife Science Module 4 Lesson 5 and 6Mae AlfaroNo ratings yet
- GENERAL BIOLOGY Q2 WEEK 6 Glycolysis and Kreb CycleDocument14 pagesGENERAL BIOLOGY Q2 WEEK 6 Glycolysis and Kreb CycleAryan Jovic DomingoNo ratings yet
- 1.2. Biomolecules CarbsLipidsProt 22-23Document102 pages1.2. Biomolecules CarbsLipidsProt 22-23Guillem DehesaNo ratings yet
- 10 Catabolism Reading ModuleDocument33 pages10 Catabolism Reading ModuleSebastian SmytheNo ratings yet
- ResultDocument15 pagesResultWei Loon100% (1)
- MetabolismDocument51 pagesMetabolismHanna YajNo ratings yet
- Briefly Summarize The Involving Steps in The ATP SynthesisDocument7 pagesBriefly Summarize The Involving Steps in The ATP SynthesisXuân ViNo ratings yet
- BioenergeticsDocument29 pagesBioenergeticskizzaymenteraNo ratings yet
- Metabolism and Metabolic Pathways in Bioprocessing Metabolism and Metabolic Pathways in BioprocessingDocument101 pagesMetabolism and Metabolic Pathways in Bioprocessing Metabolism and Metabolic Pathways in BioprocessingAhmadin BamudNo ratings yet
- CR Lab ReportDocument6 pagesCR Lab ReportslowteeNo ratings yet
- Physioex Digestion (Fin)Document12 pagesPhysioex Digestion (Fin)Tisha TuazonNo ratings yet
- CarbohydratesDocument19 pagesCarbohydratesNaina YadavvNo ratings yet
- Unit 1: Seat of Wisdom Secondary School, Trans Ekulu, EnuguDocument20 pagesUnit 1: Seat of Wisdom Secondary School, Trans Ekulu, EnuguUkwuma Michael ChijiokeNo ratings yet
- Elemental Composition of BacteriaDocument11 pagesElemental Composition of Bacteriaishmist04No ratings yet
- Carbohydrate MetabolismDocument19 pagesCarbohydrate MetabolismAbhithNo ratings yet
- The Fate of Metabolism Andmabolic PathwaysDocument25 pagesThe Fate of Metabolism Andmabolic PathwaysXuân Vi100% (1)
- MetabolismeDocument33 pagesMetabolismeNiza LailaNo ratings yet
- Session #6 SAS - Nutrition (Lecture)Document6 pagesSession #6 SAS - Nutrition (Lecture)Mariel Gwen RetorcaNo ratings yet
- Carbohydrate Distribution, Metabolism, and ExcretionDocument93 pagesCarbohydrate Distribution, Metabolism, and ExcretionPSPD - Hana Athaya NNo ratings yet
- IV.A. Nutrient Absorption in CellsDocument14 pagesIV.A. Nutrient Absorption in CellsDarwin NoolNo ratings yet
- Cellular Respiration and Fermentation (2 Files Merged) PDFDocument93 pagesCellular Respiration and Fermentation (2 Files Merged) PDFeddula ganeshNo ratings yet
- Ethanol Production by Wine FermentationDocument6 pagesEthanol Production by Wine FermentationSabyasachi DasguptaNo ratings yet
- Topic 3 Microbes and MetabolismDocument55 pagesTopic 3 Microbes and MetabolismhayyatiNo ratings yet
- Review ArticleDocument20 pagesReview ArticleAHKI ANTONETTE ABELNo ratings yet
- Grade 12 LM Physical Science 1 Module4Document21 pagesGrade 12 LM Physical Science 1 Module4ladyheart ۦۦNo ratings yet
- Lipid Metabolism Dental and PhysiotherapyDocument43 pagesLipid Metabolism Dental and PhysiotherapyNada Atef KoraitemNo ratings yet
- Chapter 8 AnswersDocument12 pagesChapter 8 Answersdaniell.baii2526No ratings yet
- Hsslive-xi-botany-RESPIRATION IN PLANTS-8Document7 pagesHsslive-xi-botany-RESPIRATION IN PLANTS-8diyakozhimalaNo ratings yet
- Glossary Key PointsDocument3 pagesGlossary Key PointsbxyrtbgyxcNo ratings yet
- Cellular Metabolism Overview of MetabolismDocument4 pagesCellular Metabolism Overview of MetabolismNathan Louis PalacioNo ratings yet
- Carbohydrate, Protein and Lipid Metabolism-B.NDocument77 pagesCarbohydrate, Protein and Lipid Metabolism-B.NBusy worldNo ratings yet
- Assignment in BiochemDocument5 pagesAssignment in BiochemEnrique FrigillanaNo ratings yet
- Module 5 Lesson 12Document13 pagesModule 5 Lesson 12John eric CatayongNo ratings yet
- Assignment No 2 Course Title: Bio Chemistry Course Code: Chem-4Document6 pagesAssignment No 2 Course Title: Bio Chemistry Course Code: Chem-4Jamila RaniNo ratings yet
- Biology Reviewer (1) - CompressedDocument61 pagesBiology Reviewer (1) - CompressedkevinNo ratings yet
- Digestive System: Faculty of Math and Science, Makassar State University EmailDocument5 pagesDigestive System: Faculty of Math and Science, Makassar State University EmailNur RahmahNo ratings yet
- AQA Biology Unit 1: Revision Notes: myrevisionnotes, #1From EverandAQA Biology Unit 1: Revision Notes: myrevisionnotes, #1Rating: 5 out of 5 stars5/5 (2)
- Nucleic AcidDocument28 pagesNucleic Acidkjgbitoon00125No ratings yet
- Carbohydrate MetabolismDocument101 pagesCarbohydrate Metabolismkjgbitoon00125No ratings yet
- Hydro MidtermDocument6 pagesHydro Midtermkjgbitoon00125No ratings yet
- Abe Law QuizDocument5 pagesAbe Law Quizkjgbitoon00125No ratings yet
- Pascal, 2011Document10 pagesPascal, 2011anaNo ratings yet
- Unit-1 Basic Concept of MetabolismDocument31 pagesUnit-1 Basic Concept of Metabolismsajidraza9821No ratings yet
- BioRxn Eng PDFDocument638 pagesBioRxn Eng PDFalabastrinoNo ratings yet
- 2012 HSC Exam ChemistryDocument40 pages2012 HSC Exam ChemistrynigerianhacksNo ratings yet
- Optimization of Xanthan Gum Fermentation Utilizing Food WasteDocument11 pagesOptimization of Xanthan Gum Fermentation Utilizing Food WasteGRD JournalsNo ratings yet
- 92.physiology of Lens and Cataractogenesis... SrisDocument92 pages92.physiology of Lens and Cataractogenesis... SrisSristi ThakurNo ratings yet
- Muscle Metabolic Adaptation To Exercise: Learning ObjectivesDocument6 pagesMuscle Metabolic Adaptation To Exercise: Learning ObjectivesUloko ChristopherNo ratings yet
- Bio-Chemistry MCQ - No AnswerDocument8 pagesBio-Chemistry MCQ - No AnswerVichhaiJackson100% (1)
- Ovber StartDocument12 pagesOvber StartHemali Sudhakar NarkhedeNo ratings yet
- Biology Concepts and Connections Campbell 6th Edition Test BankDocument25 pagesBiology Concepts and Connections Campbell 6th Edition Test BankRichard RunnelsNo ratings yet
- Soi 2020Document246 pagesSoi 2020nitinNo ratings yet
- General BiologyDocument36 pagesGeneral BiologyAdrienne CabanigNo ratings yet
- Jurol Fenol Ing 1Document7 pagesJurol Fenol Ing 1milktea segerrNo ratings yet
- PBIO211Lec FINALS 1Document26 pagesPBIO211Lec FINALS 1richard respetoNo ratings yet
- Biological Degradation of Cyanide Compounds: Stephen EbbsDocument6 pagesBiological Degradation of Cyanide Compounds: Stephen EbbsEva Pa'e ONo ratings yet
- Biochemistry 4th Edition Mathews Test Bank 1Document36 pagesBiochemistry 4th Edition Mathews Test Bank 1loribowersbwknifoyxe100% (29)
- Sas 6 1Document18 pagesSas 6 1Reizel GaasNo ratings yet
- Chapter 1: Bacterialcell Structure, Physiology, Metabolism, and GeneticsDocument19 pagesChapter 1: Bacterialcell Structure, Physiology, Metabolism, and GeneticsNico Loko100% (1)
- BIOMO303 Test and Exam Questions (Part I)Document2 pagesBIOMO303 Test and Exam Questions (Part I)AdrianNo ratings yet
- Bioethics, The Science of SurvivalDocument28 pagesBioethics, The Science of SurvivalAlejandro Andrés Crespo VillaNo ratings yet
- BioBuild Desert CrossingDocument8 pagesBioBuild Desert CrossingSubbaraju GvNo ratings yet
- ELSci Q2 Lesson 5 - Metabolic Processes Among Living ThingsDocument51 pagesELSci Q2 Lesson 5 - Metabolic Processes Among Living ThingsItsClarenceNo ratings yet
- 9 Bacterial MetabolismDocument7 pages9 Bacterial MetabolismAlphonse Rossaint SambranoNo ratings yet
- Wardlaws Perspectives in Nutrition A Functional Approach 1st Edition Byrd Bredbenner Test BankDocument38 pagesWardlaws Perspectives in Nutrition A Functional Approach 1st Edition Byrd Bredbenner Test Bankedricduyenuc1uw100% (26)
- Biomolecules Crash Course NotesDocument22 pagesBiomolecules Crash Course NotesAayush sainiNo ratings yet
- B. Tech. Biotechnology Syllabus CDFSTDocument42 pagesB. Tech. Biotechnology Syllabus CDFSTlakshitaNo ratings yet
- Lipid Metabolism VijayDocument30 pagesLipid Metabolism VijaytiffNo ratings yet




























































































Download as pdf or txt
You might also like
- Biochemistry II End of Semester Exam QuestionsDocument6 pagesBiochemistry II End of Semester Exam Questionsnima00No ratings yet
- Biochemistry MCQsDocument19 pagesBiochemistry MCQsschxzerrydawn100% (1)
- Testbank 4e CH14Document19 pagesTestbank 4e CH14somasudeshna100% (5)
- UntitledDocument14 pagesUntitledSyafrianda PanggabeanNo ratings yet
- Unit 8 Digestion and AbsorptionDocument33 pagesUnit 8 Digestion and AbsorptionGlory MuedaNo ratings yet
- Biology M5 Cellular RespirationDocument30 pagesBiology M5 Cellular RespirationpatrickkayeNo ratings yet
- Docs 2Document13 pagesDocs 2Mary Rose CuentasNo ratings yet
- 04 Energy TransformationDocument53 pages04 Energy TransformationlyleandreihendersonNo ratings yet
- Lecture 02. Common Metabolic Pathways. Biological Oxidation. Tissue Respiration. Oxidative PhosphorylationDocument53 pagesLecture 02. Common Metabolic Pathways. Biological Oxidation. Tissue Respiration. Oxidative PhosphorylationВіталій Михайлович НечипорукNo ratings yet
- MODULE - FINALS - Biochem - 1Document5 pagesMODULE - FINALS - Biochem - 1Cyril CauilanNo ratings yet
- GEN BIO RevsDocument7 pagesGEN BIO RevsacglianeNo ratings yet
- Pre Test: Prepared By: Patricia PradoDocument51 pagesPre Test: Prepared By: Patricia PradoLovely AlosadoNo ratings yet
- Module 12 PDFDocument13 pagesModule 12 PDFJohn Bernard VeloriaNo ratings yet
- Biochemistry (Metabolism) by Moses KDocument405 pagesBiochemistry (Metabolism) by Moses Khaamajansi86% (7)
- Module 8 Metabolic Pathways Final 1Document56 pagesModule 8 Metabolic Pathways Final 1Susana N peNo ratings yet
- 3 CarbohydratesDocument12 pages3 CarbohydratesconcepcionsofiakarylleNo ratings yet
- How Animals Survive Lesson 5Document30 pagesHow Animals Survive Lesson 5Ellah LaguismaNo ratings yet
- How Cells Release Energy Part 1Document14 pagesHow Cells Release Energy Part 1Jiro SarmientoNo ratings yet
- 05 F4 Biology Chap 6Document19 pages05 F4 Biology Chap 6Benjamin TeeNo ratings yet
- The Biochemistry of Digestion, Absorption and Detoxification by Prof. Dr. Hedef D. El-YassinDocument64 pagesThe Biochemistry of Digestion, Absorption and Detoxification by Prof. Dr. Hedef D. El-YassinSayhidoen CepexNo ratings yet
- Agri-501 Biochemistry 2021-2022: Doc. Regina P. ClavelDocument36 pagesAgri-501 Biochemistry 2021-2022: Doc. Regina P. ClavelIvan Jhon AnamNo ratings yet
- Biology The Essentials 1st Edition Hoefnagels Solutions Manual DownloadDocument15 pagesBiology The Essentials 1st Edition Hoefnagels Solutions Manual DownloadGraham Hissem100% (26)
- Carbohydrates Part 2 - MetabolismsDocument60 pagesCarbohydrates Part 2 - MetabolismsNikka Mia AbadiesNo ratings yet
- Metabolic ProcessesDocument6 pagesMetabolic ProcessesMark Andrian PontanalNo ratings yet
- Last Cellmolecule ReportDocument10 pagesLast Cellmolecule Reportapi-638362671No ratings yet
- Life Science Module 4 Lesson 5 and 6Document38 pagesLife Science Module 4 Lesson 5 and 6Mae AlfaroNo ratings yet
- GENERAL BIOLOGY Q2 WEEK 6 Glycolysis and Kreb CycleDocument14 pagesGENERAL BIOLOGY Q2 WEEK 6 Glycolysis and Kreb CycleAryan Jovic DomingoNo ratings yet
- 1.2. Biomolecules CarbsLipidsProt 22-23Document102 pages1.2. Biomolecules CarbsLipidsProt 22-23Guillem DehesaNo ratings yet
- 10 Catabolism Reading ModuleDocument33 pages10 Catabolism Reading ModuleSebastian SmytheNo ratings yet
- ResultDocument15 pagesResultWei Loon100% (1)
- MetabolismDocument51 pagesMetabolismHanna YajNo ratings yet
- Briefly Summarize The Involving Steps in The ATP SynthesisDocument7 pagesBriefly Summarize The Involving Steps in The ATP SynthesisXuân ViNo ratings yet
- BioenergeticsDocument29 pagesBioenergeticskizzaymenteraNo ratings yet
- Metabolism and Metabolic Pathways in Bioprocessing Metabolism and Metabolic Pathways in BioprocessingDocument101 pagesMetabolism and Metabolic Pathways in Bioprocessing Metabolism and Metabolic Pathways in BioprocessingAhmadin BamudNo ratings yet
- CR Lab ReportDocument6 pagesCR Lab ReportslowteeNo ratings yet
- Physioex Digestion (Fin)Document12 pagesPhysioex Digestion (Fin)Tisha TuazonNo ratings yet
- CarbohydratesDocument19 pagesCarbohydratesNaina YadavvNo ratings yet
- Unit 1: Seat of Wisdom Secondary School, Trans Ekulu, EnuguDocument20 pagesUnit 1: Seat of Wisdom Secondary School, Trans Ekulu, EnuguUkwuma Michael ChijiokeNo ratings yet
- Elemental Composition of BacteriaDocument11 pagesElemental Composition of Bacteriaishmist04No ratings yet
- Carbohydrate MetabolismDocument19 pagesCarbohydrate MetabolismAbhithNo ratings yet
- The Fate of Metabolism Andmabolic PathwaysDocument25 pagesThe Fate of Metabolism Andmabolic PathwaysXuân Vi100% (1)
- MetabolismeDocument33 pagesMetabolismeNiza LailaNo ratings yet
- Session #6 SAS - Nutrition (Lecture)Document6 pagesSession #6 SAS - Nutrition (Lecture)Mariel Gwen RetorcaNo ratings yet
- Carbohydrate Distribution, Metabolism, and ExcretionDocument93 pagesCarbohydrate Distribution, Metabolism, and ExcretionPSPD - Hana Athaya NNo ratings yet
- IV.A. Nutrient Absorption in CellsDocument14 pagesIV.A. Nutrient Absorption in CellsDarwin NoolNo ratings yet
- Cellular Respiration and Fermentation (2 Files Merged) PDFDocument93 pagesCellular Respiration and Fermentation (2 Files Merged) PDFeddula ganeshNo ratings yet
- Ethanol Production by Wine FermentationDocument6 pagesEthanol Production by Wine FermentationSabyasachi DasguptaNo ratings yet
- Topic 3 Microbes and MetabolismDocument55 pagesTopic 3 Microbes and MetabolismhayyatiNo ratings yet
- Review ArticleDocument20 pagesReview ArticleAHKI ANTONETTE ABELNo ratings yet
- Grade 12 LM Physical Science 1 Module4Document21 pagesGrade 12 LM Physical Science 1 Module4ladyheart ۦۦNo ratings yet
- Lipid Metabolism Dental and PhysiotherapyDocument43 pagesLipid Metabolism Dental and PhysiotherapyNada Atef KoraitemNo ratings yet
- Chapter 8 AnswersDocument12 pagesChapter 8 Answersdaniell.baii2526No ratings yet
- Hsslive-xi-botany-RESPIRATION IN PLANTS-8Document7 pagesHsslive-xi-botany-RESPIRATION IN PLANTS-8diyakozhimalaNo ratings yet
- Glossary Key PointsDocument3 pagesGlossary Key PointsbxyrtbgyxcNo ratings yet
- Cellular Metabolism Overview of MetabolismDocument4 pagesCellular Metabolism Overview of MetabolismNathan Louis PalacioNo ratings yet
- Carbohydrate, Protein and Lipid Metabolism-B.NDocument77 pagesCarbohydrate, Protein and Lipid Metabolism-B.NBusy worldNo ratings yet
- Assignment in BiochemDocument5 pagesAssignment in BiochemEnrique FrigillanaNo ratings yet
- Module 5 Lesson 12Document13 pagesModule 5 Lesson 12John eric CatayongNo ratings yet
- Assignment No 2 Course Title: Bio Chemistry Course Code: Chem-4Document6 pagesAssignment No 2 Course Title: Bio Chemistry Course Code: Chem-4Jamila RaniNo ratings yet
- Biology Reviewer (1) - CompressedDocument61 pagesBiology Reviewer (1) - CompressedkevinNo ratings yet
- Digestive System: Faculty of Math and Science, Makassar State University EmailDocument5 pagesDigestive System: Faculty of Math and Science, Makassar State University EmailNur RahmahNo ratings yet
- AQA Biology Unit 1: Revision Notes: myrevisionnotes, #1From EverandAQA Biology Unit 1: Revision Notes: myrevisionnotes, #1Rating: 5 out of 5 stars5/5 (2)
- Nucleic AcidDocument28 pagesNucleic Acidkjgbitoon00125No ratings yet
- Carbohydrate MetabolismDocument101 pagesCarbohydrate Metabolismkjgbitoon00125No ratings yet
- Hydro MidtermDocument6 pagesHydro Midtermkjgbitoon00125No ratings yet
- Abe Law QuizDocument5 pagesAbe Law Quizkjgbitoon00125No ratings yet
- Pascal, 2011Document10 pagesPascal, 2011anaNo ratings yet
- Unit-1 Basic Concept of MetabolismDocument31 pagesUnit-1 Basic Concept of Metabolismsajidraza9821No ratings yet
- BioRxn Eng PDFDocument638 pagesBioRxn Eng PDFalabastrinoNo ratings yet
- 2012 HSC Exam ChemistryDocument40 pages2012 HSC Exam ChemistrynigerianhacksNo ratings yet
- Optimization of Xanthan Gum Fermentation Utilizing Food WasteDocument11 pagesOptimization of Xanthan Gum Fermentation Utilizing Food WasteGRD JournalsNo ratings yet
- 92.physiology of Lens and Cataractogenesis... SrisDocument92 pages92.physiology of Lens and Cataractogenesis... SrisSristi ThakurNo ratings yet
- Muscle Metabolic Adaptation To Exercise: Learning ObjectivesDocument6 pagesMuscle Metabolic Adaptation To Exercise: Learning ObjectivesUloko ChristopherNo ratings yet
- Bio-Chemistry MCQ - No AnswerDocument8 pagesBio-Chemistry MCQ - No AnswerVichhaiJackson100% (1)
- Ovber StartDocument12 pagesOvber StartHemali Sudhakar NarkhedeNo ratings yet
- Biology Concepts and Connections Campbell 6th Edition Test BankDocument25 pagesBiology Concepts and Connections Campbell 6th Edition Test BankRichard RunnelsNo ratings yet
- Soi 2020Document246 pagesSoi 2020nitinNo ratings yet
- General BiologyDocument36 pagesGeneral BiologyAdrienne CabanigNo ratings yet
- Jurol Fenol Ing 1Document7 pagesJurol Fenol Ing 1milktea segerrNo ratings yet
- PBIO211Lec FINALS 1Document26 pagesPBIO211Lec FINALS 1richard respetoNo ratings yet
- Biological Degradation of Cyanide Compounds: Stephen EbbsDocument6 pagesBiological Degradation of Cyanide Compounds: Stephen EbbsEva Pa'e ONo ratings yet
- Biochemistry 4th Edition Mathews Test Bank 1Document36 pagesBiochemistry 4th Edition Mathews Test Bank 1loribowersbwknifoyxe100% (29)
- Sas 6 1Document18 pagesSas 6 1Reizel GaasNo ratings yet
- Chapter 1: Bacterialcell Structure, Physiology, Metabolism, and GeneticsDocument19 pagesChapter 1: Bacterialcell Structure, Physiology, Metabolism, and GeneticsNico Loko100% (1)
- BIOMO303 Test and Exam Questions (Part I)Document2 pagesBIOMO303 Test and Exam Questions (Part I)AdrianNo ratings yet
- Bioethics, The Science of SurvivalDocument28 pagesBioethics, The Science of SurvivalAlejandro Andrés Crespo VillaNo ratings yet
- BioBuild Desert CrossingDocument8 pagesBioBuild Desert CrossingSubbaraju GvNo ratings yet
- ELSci Q2 Lesson 5 - Metabolic Processes Among Living ThingsDocument51 pagesELSci Q2 Lesson 5 - Metabolic Processes Among Living ThingsItsClarenceNo ratings yet
- 9 Bacterial MetabolismDocument7 pages9 Bacterial MetabolismAlphonse Rossaint SambranoNo ratings yet
- Wardlaws Perspectives in Nutrition A Functional Approach 1st Edition Byrd Bredbenner Test BankDocument38 pagesWardlaws Perspectives in Nutrition A Functional Approach 1st Edition Byrd Bredbenner Test Bankedricduyenuc1uw100% (26)
- Biomolecules Crash Course NotesDocument22 pagesBiomolecules Crash Course NotesAayush sainiNo ratings yet
- B. Tech. Biotechnology Syllabus CDFSTDocument42 pagesB. Tech. Biotechnology Syllabus CDFSTlakshitaNo ratings yet
- Lipid Metabolism VijayDocument30 pagesLipid Metabolism VijaytiffNo ratings yet