Download as pdf or txt
You might also like
- Wong's Essentials of Pediatric Nursing, 10th Edition - Test BankDocument44 pagesWong's Essentials of Pediatric Nursing, 10th Edition - Test BankalishcathrinNo ratings yet
- Deagel Analysis UpdatedDocument7 pagesDeagel Analysis UpdatedAll News PipelineNo ratings yet
- tmp37AD TMPDocument11 pagestmp37AD TMPFrontiersNo ratings yet
- Cancer associated fibroblast derived SLIT2 drives gastric cancer cell metastasis by activating NEK9Document13 pagesCancer associated fibroblast derived SLIT2 drives gastric cancer cell metastasis by activating NEK9nasirlayla0No ratings yet
- KDM5B MacrophageDocument14 pagesKDM5B MacrophageMaurício DallowayNo ratings yet
- LCH TargetDocument5 pagesLCH TargetSeham GoharNo ratings yet
- Cytoglobin Inhibits Migration Through PI3K/AKT/mTOR Pathway in Fibroblast CellsDocument10 pagesCytoglobin Inhibits Migration Through PI3K/AKT/mTOR Pathway in Fibroblast CellsDilay CelikNo ratings yet
- Immunometabolism in The Single-Cell Era: PerspectiveDocument16 pagesImmunometabolism in The Single-Cell Era: PerspectiveyasbiloNo ratings yet
- 2022 International Immunopharmacology Q1RamírezDocument15 pages2022 International Immunopharmacology Q1RamírezAndres RamirezNo ratings yet
- International ImmunopharmacologyDocument15 pagesInternational ImmunopharmacologyAndres RamirezNo ratings yet
- CSF3R 6Document7 pagesCSF3R 6AndikhaNo ratings yet
- Low Molecular Weight Hyaluronan Mediated CD44 Dependent InductionDocument7 pagesLow Molecular Weight Hyaluronan Mediated CD44 Dependent Inductiond.aouameurNo ratings yet
- SM22α suppresses cytokine-induced inflammation and the transcription of NF-κB inducing kinase (Nik) by modulating SRF transcriptional activity in vascular smooth muscle cellsDocument11 pagesSM22α suppresses cytokine-induced inflammation and the transcription of NF-κB inducing kinase (Nik) by modulating SRF transcriptional activity in vascular smooth muscle cellsaneerajapNo ratings yet
- European Journal of PharmacologyDocument9 pagesEuropean Journal of PharmacologyDumitru RadulescuNo ratings yet
- Chemically Induced Reprogramming of Somatic Cells To Pluripotent Stem Cells and Neural CellsDocument13 pagesChemically Induced Reprogramming of Somatic Cells To Pluripotent Stem Cells and Neural CellsnembutalNo ratings yet
- MEK Inhibitors Activate WNT Signalling and Induce Stem Cell Plasticity in Colorectal CancerDocument17 pagesMEK Inhibitors Activate WNT Signalling and Induce Stem Cell Plasticity in Colorectal CancerYuqing LiangNo ratings yet
- Phytomedicine: Original ArticleDocument9 pagesPhytomedicine: Original ArticleAnjali MohanNo ratings yet
- Macrophage Colony-Stimulating Factor and Cancer A ReviewDocument10 pagesMacrophage Colony-Stimulating Factor and Cancer A ReviewCristian Gutiérrez VeraNo ratings yet
- Sistem Informasi Kesehatan (Leukimia)Document22 pagesSistem Informasi Kesehatan (Leukimia)YuliaNo ratings yet
- Akabl I 2018Document9 pagesAkabl I 2018taoufik akabliNo ratings yet
- 2010 Articulo TGF-b1 and ROS Tobar Mol Cell Biochem 2010-LibreDocument8 pages2010 Articulo TGF-b1 and ROS Tobar Mol Cell Biochem 2010-LibretirtasetiawanNo ratings yet
- At The Crossroads of Lipid Metabolism and InflammationDocument7 pagesAt The Crossroads of Lipid Metabolism and InflammationChow Sing SingNo ratings yet
- Sanz Moreno2012Document3 pagesSanz Moreno2012Ernesto MoralesNo ratings yet
- Bk-Virus Infections A Literature ReviewDocument5 pagesBk-Virus Infections A Literature Reviewfvffv0x7100% (1)
- Nowag - 2023 - J Cosmet Dermatol. Biostimulating Fillers and Induction of Inflammatory PathwaysDocument8 pagesNowag - 2023 - J Cosmet Dermatol. Biostimulating Fillers and Induction of Inflammatory PathwaysdracelestedimatteoNo ratings yet
- Trudel 1988Document10 pagesTrudel 1988nickiNo ratings yet
- Hsiah2 Is A New Vav Binding Protein Which Inhibits Vav-Mediated Signaling PathwaysDocument10 pagesHsiah2 Is A New Vav Binding Protein Which Inhibits Vav-Mediated Signaling PathwaysFernando JarixNo ratings yet
- Effect of Spautin 1 On Dengue JVI 2013Document10 pagesEffect of Spautin 1 On Dengue JVI 2013Kudelko MatNo ratings yet
- NMV IsolationDocument10 pagesNMV IsolationManovriti ThakurNo ratings yet
- Much More Than M1 and M2 Macrophages, There Are Also CD169 and TCR MacrophagesDocument15 pagesMuch More Than M1 and M2 Macrophages, There Are Also CD169 and TCR MacrophagesHolman FuyaNo ratings yet
- 2019jarell CFD Celldelivery mRNA VorticesDocument11 pages2019jarell CFD Celldelivery mRNA VorticesSai Santhosh ManepallyNo ratings yet
- Tumor Cell Pseudopodial ProtrusionsDocument10 pagesTumor Cell Pseudopodial Protrusionsvanhuynh2812No ratings yet
- 1953 FullDocument10 pages1953 FullboboyoyotNo ratings yet
- S74 Poster Abstracts: Table 1Document1 pageS74 Poster Abstracts: Table 1HanaNo ratings yet
- Mecanismos Que Regulan La Actividad Del Receptor de Quimiocinas 2011Document11 pagesMecanismos Que Regulan La Actividad Del Receptor de Quimiocinas 2011oscarbio2009No ratings yet
- Genome Maintenance Meets MechanobiologyDocument22 pagesGenome Maintenance Meets MechanobiologyMARIMORRIS 006No ratings yet
- 2014 GorardiDocument16 pages2014 GorardiAtrocitus RedNo ratings yet
- Silibinin Inhibits Proliferation and Migration of Human Hepatic Stellate LX-2 CellsDocument3 pagesSilibinin Inhibits Proliferation and Migration of Human Hepatic Stellate LX-2 CellsSILVIA BAQUEIRONo ratings yet
- MicroDocument11 pagesMicroParijat BanerjeeNo ratings yet
- Do We Know JackDocument20 pagesDo We Know JackAbdul-Ganiyu MannirNo ratings yet
- Cold Spring Harb Perspect Biol-2015-Sokol-A016303Document20 pagesCold Spring Harb Perspect Biol-2015-Sokol-A016303王鑫No ratings yet
- Temi 11 2065935Document12 pagesTemi 11 2065935Robert StryjakNo ratings yet
- Godwin 2013 Macrophages Are Required For Adult Salamander Limb RegenerationDocument6 pagesGodwin 2013 Macrophages Are Required For Adult Salamander Limb Regenerationafh230385No ratings yet
- The Interplay Between - TspaceDocument35 pagesThe Interplay Between - TspaceArooNo ratings yet
- Cells 10 01540Document20 pagesCells 10 01540klikpeneliti05No ratings yet
- A Chemical Biology Screen Identifies Glucocorticoids That Regulate C - 2007 - BLDocument8 pagesA Chemical Biology Screen Identifies Glucocorticoids That Regulate C - 2007 - BLTareeqanwar MohammedNo ratings yet
- 19S-19-Acetoxychavicol Acetate Isolated From Alpinia Galanga Inhibits Human Immunodeficiency Virus Type 1 Replication by Blocking Rev TransportDocument7 pages19S-19-Acetoxychavicol Acetate Isolated From Alpinia Galanga Inhibits Human Immunodeficiency Virus Type 1 Replication by Blocking Rev TransportVictoria RookerNo ratings yet
- DOC13Document7 pagesDOC13begoñaNo ratings yet
- Evasin-Displaying Lactic Acid Bacteria Bind DifferDocument12 pagesEvasin-Displaying Lactic Acid Bacteria Bind DifferGregorius HocevarNo ratings yet
- Pasq 2004 BloodDocument8 pagesPasq 2004 BloodlillareinigerNo ratings yet
- BMC Evolutionary Biology: Phylogenetic Diversity of Stress Signalling Pathways in FungiDocument18 pagesBMC Evolutionary Biology: Phylogenetic Diversity of Stress Signalling Pathways in Fungisarns456No ratings yet
- 0300060520925693Document11 pages0300060520925693Narjes Sharif AskariNo ratings yet
- Inter-Cellular Nanovesicle Mediated MicroRNA TransferDocument19 pagesInter-Cellular Nanovesicle Mediated MicroRNA Transferanon_579968442No ratings yet
- Resveratrol Promotes Autophagic Cell Death in Chronic Myelogenous Leukemia Cells Via JNK-Mediated p62/SQSTM1 Expression and AMPK ActivationDocument11 pagesResveratrol Promotes Autophagic Cell Death in Chronic Myelogenous Leukemia Cells Via JNK-Mediated p62/SQSTM1 Expression and AMPK Activationhorsepower735No ratings yet
- 3 PBDocument12 pages3 PBDumitruNo ratings yet
- Exosomes Derived From Mir-26A-Modified Mscs Promote Axonal Regeneration Via The Pten/Akt/Mtor Pathway Following Spinal Cord InjuryDocument15 pagesExosomes Derived From Mir-26A-Modified Mscs Promote Axonal Regeneration Via The Pten/Akt/Mtor Pathway Following Spinal Cord Injuryelif.onsozNo ratings yet
- Omcl2021 9068850Document31 pagesOmcl2021 9068850Heloisa PessoaNo ratings yet
- Nuclear Factor Kappa B (NF B) A Key Factor Involved in Hypoxia-Induced Chemoresistance and Stemness of Breast Cancer CellsDocument10 pagesNuclear Factor Kappa B (NF B) A Key Factor Involved in Hypoxia-Induced Chemoresistance and Stemness of Breast Cancer CellsInternational Journal of Advance Study and Research WorkNo ratings yet
- Growth Factor Receptor Signaling Inhibition Prevents Sars-Cov-2 ReplicationDocument28 pagesGrowth Factor Receptor Signaling Inhibition Prevents Sars-Cov-2 ReplicationBf LiawNo ratings yet
- Immune Cell Metabolism and Metabolic ReprogrammingDocument13 pagesImmune Cell Metabolism and Metabolic ReprogrammingFlávia PampolhaNo ratings yet
- LAK and DCsDocument9 pagesLAK and DCsFabiane ColunnaNo ratings yet
- Cancer Gene Therapy by Viral and Non-viral VectorsFrom EverandCancer Gene Therapy by Viral and Non-viral VectorsMalcolm BrennerNo ratings yet
- Idt 17 2Document44 pagesIdt 17 2anhchangcodon88No ratings yet
- Full download Dermatology Essentials 2nd Edition Jean L. Bolognia file pdf all chapter on 2024Document44 pagesFull download Dermatology Essentials 2nd Edition Jean L. Bolognia file pdf all chapter on 2024sabbertsunkeNo ratings yet
- Jurnal Upn FadhliDocument15 pagesJurnal Upn FadhliFadhli Dzil IkramNo ratings yet
- Florence Nightingale Theory: She Emphasizes Five Essential Point in Securing Health of HouseDocument2 pagesFlorence Nightingale Theory: She Emphasizes Five Essential Point in Securing Health of HouseAkshata BansodeNo ratings yet
- Periodontics I/Lecture3 Etiology of Periodontal Diseases: Dental PlaqueDocument12 pagesPeriodontics I/Lecture3 Etiology of Periodontal Diseases: Dental Plaqueربيد احمد مثنى يحيى كلية طب الاسنان - جامعة عدنNo ratings yet
- GeneticTestPdf 6538 PDFDocument21 pagesGeneticTestPdf 6538 PDFMarianaBBazanNo ratings yet
- ACOG Fetal Maturity Guidelines PDFDocument8 pagesACOG Fetal Maturity Guidelines PDFinayahsyNo ratings yet
- Jurnal Massage For Infantile ColicDocument10 pagesJurnal Massage For Infantile ColicrambuubeNo ratings yet
- Anesthesia Management of Pregnant With HELLP Syndrome With Fetal Intrauterine Exitus (#1123666) - 2457074Document3 pagesAnesthesia Management of Pregnant With HELLP Syndrome With Fetal Intrauterine Exitus (#1123666) - 2457074SALMA HANINANo ratings yet
- Linder2019 PDFDocument3 pagesLinder2019 PDFDian Putri NingsihNo ratings yet
- Mengapa Biosecurity Menjadi Penting Pada Laboratorium Penyakit Infeksi ?Document10 pagesMengapa Biosecurity Menjadi Penting Pada Laboratorium Penyakit Infeksi ?AndyNo ratings yet
- Cefazolin Drug StudyDocument2 pagesCefazolin Drug StudyAj GoNo ratings yet
- To Study Oxalate Ion in GuavaDocument15 pagesTo Study Oxalate Ion in Guavasureshkumar ChemNo ratings yet
- Trastornos de La Conducta AlimentariaDocument10 pagesTrastornos de La Conducta AlimentariaAlejandra CarrilloNo ratings yet
- Pediatrics - Neonatal ICUDocument2 pagesPediatrics - Neonatal ICUJasmine KangNo ratings yet
- Management of Prolonged Second Stage of Labor: Sanjay Singh, Uttara Aiyer Kohli, Shakti VardhanDocument5 pagesManagement of Prolonged Second Stage of Labor: Sanjay Singh, Uttara Aiyer Kohli, Shakti VardhansheilaNo ratings yet
- The Effect of Manual Therapy and Neuroplasticity Education On Chronic Low Back Pain: A Randomized Clinical TrialDocument10 pagesThe Effect of Manual Therapy and Neuroplasticity Education On Chronic Low Back Pain: A Randomized Clinical TrialVitor SilvaNo ratings yet
- NSCRP Tool ComponentsDocument3 pagesNSCRP Tool ComponentsCelyne UlarteNo ratings yet
- Bladder Catheterisation PDFDocument12 pagesBladder Catheterisation PDFSonish KhatriNo ratings yet
- Raynaud's Disease & Buerger's Disease: Submitted ToDocument11 pagesRaynaud's Disease & Buerger's Disease: Submitted ToKathleen Leana Viray JeanjaquetNo ratings yet
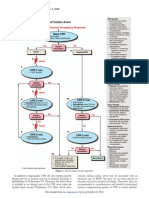
- ACLS AlgorithmsDocument4 pagesACLS Algorithmsmonickams100% (1)
- Drug Study For Prednisone Case INPDocument2 pagesDrug Study For Prednisone Case INPChristina Barroga100% (4)
- VARICOSE VEINS .BY DR BasharatDocument94 pagesVARICOSE VEINS .BY DR BasharatMohammad NomanNo ratings yet
- Scapulohumeral PeriarthritisDocument29 pagesScapulohumeral PeriarthritisMárcia PatríciaNo ratings yet
- Evidence-Based Practice Project: Diabetes Diet Management: Stephanie Green NUR - 590Document20 pagesEvidence-Based Practice Project: Diabetes Diet Management: Stephanie Green NUR - 590dennis ndegeNo ratings yet
- Krisis Manusia Modern Dan Pendidikan IslamDocument17 pagesKrisis Manusia Modern Dan Pendidikan IslamSyifa Nuraini RhamadhaniNo ratings yet
- Isolation and Detection of Staphylococcus Aureus in Foods: Shannen Krishna S. Sena Group 2, Sec. B-1LDocument13 pagesIsolation and Detection of Staphylococcus Aureus in Foods: Shannen Krishna S. Sena Group 2, Sec. B-1LShannen SenaNo ratings yet
- Vitamin D Deficiency Is Associated With Increased Length of Stay After Acute Burn Injury: A Multicenter AnalysisDocument5 pagesVitamin D Deficiency Is Associated With Increased Length of Stay After Acute Burn Injury: A Multicenter Analysisdds (TMU)No ratings yet