Download as pdf or txt
You might also like
- Enzyme Kinetics Lab ReportDocument20 pagesEnzyme Kinetics Lab ReportYasmin Lau100% (2)
- Summary & Study Guide - A Crack in Creation: Gene Editing and the Unthinkable Power to Control EvolutionFrom EverandSummary & Study Guide - A Crack in Creation: Gene Editing and the Unthinkable Power to Control EvolutionNo ratings yet
- Examen Traductor Oficial Colombia Modelo1 Ingles EscritoDocument6 pagesExamen Traductor Oficial Colombia Modelo1 Ingles EscritoAlejandro LandinezNo ratings yet
- Dna RepairDocument20 pagesDna RepairEaron Van JaboliNo ratings yet
- L20 - DNA Repair & RecombinationDocument41 pagesL20 - DNA Repair & RecombinationLeroy ChengNo ratings yet
- 25.2 DNA Repairv2Document30 pages25.2 DNA Repairv2ngguanfangNo ratings yet
- DNA Repair Mechanisms-1 PDFDocument21 pagesDNA Repair Mechanisms-1 PDFMehak preetNo ratings yet
- Radiobiology Lecture 3 Cellular EffectsDocument29 pagesRadiobiology Lecture 3 Cellular EffectsMankit LoNo ratings yet
- 11 03 2006 MCB Repair 1Document80 pages11 03 2006 MCB Repair 1api-3696530No ratings yet
- Exchange of Genetic MaterialDocument32 pagesExchange of Genetic MaterialMonisha SamuelNo ratings yet
- Dna Damage & RepairDocument12 pagesDna Damage & RepairDr Abhinav Manish MDNo ratings yet
- Genetic Material-DNA: 6 November 2003 Reading:The Cell Chapter 5, Pages: 192-201Document38 pagesGenetic Material-DNA: 6 November 2003 Reading:The Cell Chapter 5, Pages: 192-201Laura VgNo ratings yet
- DNA Damage Repair MechanismsDocument40 pagesDNA Damage Repair MechanismspriannaNo ratings yet
- RizalDocument25 pagesRizalPhia LorenNo ratings yet
- Introduction To Gene AnatomyDocument31 pagesIntroduction To Gene AnatomyHassan.shehri100% (1)
- DNA Damage and Tolerance Lecture NotesDocument5 pagesDNA Damage and Tolerance Lecture NotesellieNo ratings yet
- Genetic RecombinationDocument60 pagesGenetic RecombinationJeevan BasnyatNo ratings yet
- Mekanisme Replikasi, Repair DNA Dan Rekombinan DNADocument50 pagesMekanisme Replikasi, Repair DNA Dan Rekombinan DNAnolaNo ratings yet
- DNA Damage Repair MechanismDocument7 pagesDNA Damage Repair MechanismSarmistha NayakNo ratings yet
- 2 2update PDFDocument41 pages2 2update PDFJohnnyNo ratings yet
- Mbii - l15 - Dna RepairDocument6 pagesMbii - l15 - Dna RepairMiles NsgNo ratings yet
- Dna RepairDocument8 pagesDna RepairThrupthiNo ratings yet
- CH 08Document101 pagesCH 08dolt2111No ratings yet
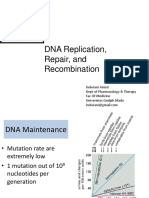
- DNA Replication, Repair, and RecombinationDocument114 pagesDNA Replication, Repair, and Recombinationviola septinaNo ratings yet
- ReplicationDocument58 pagesReplicationBillie LunaNo ratings yet
- Genetic RecombinationDocument55 pagesGenetic RecombinationRiyaniMartyasariNo ratings yet
- DNA RecombinationDocument25 pagesDNA RecombinationNedhal Mahmoud KaleefahNo ratings yet
- DNA Damage and RepairDocument24 pagesDNA Damage and Repairrajya6895No ratings yet
- Dna Replicatin and RepairDocument80 pagesDna Replicatin and RepairAndualemNo ratings yet
- DNA Replication and RepairDocument13 pagesDNA Replication and RepairViswadeep DasNo ratings yet
- Please Switch Off Your MobilesDocument28 pagesPlease Switch Off Your MobilesSadia MushtaqNo ratings yet
- Chromosome Structure and MutationDocument26 pagesChromosome Structure and MutationNikoh Anthony EwayanNo ratings yet
- DNA Replication, Repair, and RecombinationDocument92 pagesDNA Replication, Repair, and RecombinationJagdesh SajnaniNo ratings yet
- The Dna: Structure and ReplicationDocument43 pagesThe Dna: Structure and Replicationmax2711100% (1)
- DNA Rep AirDocument23 pagesDNA Rep AirTanvir AhmedNo ratings yet
- Genetic Engineering: Aliyu, HabibuDocument32 pagesGenetic Engineering: Aliyu, HabibuAkinsanmi TaiwoNo ratings yet
- Dna Repair and Cancer GoodDocument40 pagesDna Repair and Cancer GooddwNo ratings yet
- Bio 324Document4 pagesBio 324muhsinNo ratings yet
- Clase 6Document58 pagesClase 6MATEO SEBASTIAN RODRIGUEZ TORRESNo ratings yet
- HOMOLOGOUS AND NON HOMOLOGOUS RECOMBINATION by Preeti Saini 21mslsmm06Document7 pagesHOMOLOGOUS AND NON HOMOLOGOUS RECOMBINATION by Preeti Saini 21mslsmm06Preeti SainiNo ratings yet
- II ME NRP B SS 27. Biotechnology-Principles and Processes, Biotechnology - ApplicationsDocument8 pagesII ME NRP B SS 27. Biotechnology-Principles and Processes, Biotechnology - ApplicationsAmit RavindhraNo ratings yet
- Dna Damage and RepairDocument23 pagesDna Damage and RepairLathifa FauziaNo ratings yet
- Chapter20 1Document43 pagesChapter20 1as01271801497No ratings yet
- Recombinant DNA Technology: Common General Cloning StrategyDocument42 pagesRecombinant DNA Technology: Common General Cloning Strategyhugo200887No ratings yet
- Introduction To Gene Cloning: Section-ADocument48 pagesIntroduction To Gene Cloning: Section-AzxcNo ratings yet
- Genetic RecombinationDocument134 pagesGenetic RecombinationBijayaKumarUpretyNo ratings yet
- Aula BMol 2Document35 pagesAula BMol 2Diogo MadalenoNo ratings yet
- CH. 19 Replication, Repair and Recombination-2Document69 pagesCH. 19 Replication, Repair and Recombination-2IffatnazNo ratings yet
- AssignmentDocument5 pagesAssignmentSharonNo ratings yet
- DNA RepairDocument98 pagesDNA Repairapi-3700537100% (1)
- Introduction To Bacterial GeneticsDocument24 pagesIntroduction To Bacterial GeneticseyoelNo ratings yet
- 5 - Introduction To Molecular PatholgoyDocument99 pages5 - Introduction To Molecular Patholgoysara.madkour99No ratings yet
- BT 301 Short Question Mid TermDocument11 pagesBT 301 Short Question Mid TermAlishba khan100% (1)
- Lecture 26Document6 pagesLecture 26akhilakj117No ratings yet
- Gene Cloning: Dr. Sandeep Agrawal MD Senior Resident & PHD Scholar Department of Biochemistry Aiims, New DelhiDocument64 pagesGene Cloning: Dr. Sandeep Agrawal MD Senior Resident & PHD Scholar Department of Biochemistry Aiims, New Delhimahi mridulaNo ratings yet
- Oeq 1Document5 pagesOeq 1Sara JosephNo ratings yet
- DNA Replication Repair Recombination-1Document32 pagesDNA Replication Repair Recombination-1Mugo JamesNo ratings yet
- III. Molecular Genetics B Landars GroupDocument37 pagesIII. Molecular Genetics B Landars GroupMa.charlina LozadaNo ratings yet
- Lecture 5 DNA Mutation and DNA Repair-2Document65 pagesLecture 5 DNA Mutation and DNA Repair-2Constance WongNo ratings yet
- Gene Editing, Epigenetic, Cloning and TherapyFrom EverandGene Editing, Epigenetic, Cloning and TherapyRating: 4 out of 5 stars4/5 (1)
- 9700 w02 QP 4 PDFDocument12 pages9700 w02 QP 4 PDFY. PurwandariNo ratings yet
- Proteinase K, RNase - DNase Free - GoldBioDocument7 pagesProteinase K, RNase - DNase Free - GoldBioGerman PeñaNo ratings yet
- Characterization of The Human Sperm Centrosome and Its Role in (In) FertilityDocument288 pagesCharacterization of The Human Sperm Centrosome and Its Role in (In) FertilityFrogmanNo ratings yet
- 2,4-D Transport and Herbicide Resistance in WeedsDocument3 pages2,4-D Transport and Herbicide Resistance in WeedsTrần Bảo ThắngNo ratings yet
- Lab 4 Drawing of A Cross Section of A Ligustrum Plant Cell Under A Light MicroscopeDocument4 pagesLab 4 Drawing of A Cross Section of A Ligustrum Plant Cell Under A Light MicroscopeMichael TimsonNo ratings yet
- Developmental BiologyDocument28 pagesDevelopmental Biologyعمر بن عليNo ratings yet
- Hormones and Reality John S Torday Full ChapterDocument67 pagesHormones and Reality John S Torday Full Chapterjoseph.borton614100% (13)
- Textbook of Diagnostic Microbiology 7 Ed Edition Connie R Mahon Online Ebook Texxtbook Full Chapter PDFDocument69 pagesTextbook of Diagnostic Microbiology 7 Ed Edition Connie R Mahon Online Ebook Texxtbook Full Chapter PDFgene.pastor330100% (6)
- Procedures: Powerpoint Presentation, Worksheet Rabago, L M. Et Al, 2019:functional Biology, 2 Time 10 MinsDocument7 pagesProcedures: Powerpoint Presentation, Worksheet Rabago, L M. Et Al, 2019:functional Biology, 2 Time 10 Minsmichael BatungbakalNo ratings yet
- Module 2Document91 pagesModule 2Pan DemicNo ratings yet
- Xin Zhong Secondary School: School Year 2020 - 2021 Biology Worksheet Semester 1 - Grade 11 Nucleic AcidDocument6 pagesXin Zhong Secondary School: School Year 2020 - 2021 Biology Worksheet Semester 1 - Grade 11 Nucleic AcidNathan SaputroNo ratings yet
- Gen Bio3Document46 pagesGen Bio3Lorena RonquilloNo ratings yet
- Paterson Et Al. (2012) - NatureDocument6 pagesPaterson Et Al. (2012) - NatureAna Luiza Atella de FreitasNo ratings yet
- Full Download Test Bank For Evolutionary Psychology The New Science of The Mind 4 e 4th Edition David Buss PDF Full ChapterDocument36 pagesFull Download Test Bank For Evolutionary Psychology The New Science of The Mind 4 e 4th Edition David Buss PDF Full Chapterrainbowenergy.t6op100% (21)
- Biotech-Genetic Sci. TimelineDocument4 pagesBiotech-Genetic Sci. TimelinedreyyNo ratings yet
- MilliwaysDocument12 pagesMilliwaysTurtle ArtNo ratings yet
- Genetics A Conceptual Approach 5th Edition Pierce Solutions Manual 1Document22 pagesGenetics A Conceptual Approach 5th Edition Pierce Solutions Manual 1john100% (48)
- Punnett SQ Intro PracticeDocument17 pagesPunnett SQ Intro PracticeAkshata KNo ratings yet
- A Level Biology Topic 1 AssessmentDocument15 pagesA Level Biology Topic 1 AssessmentgsapkaiteNo ratings yet
- Genetic Analysis An Integrative Approach 1st Edition Sanders Test BankDocument12 pagesGenetic Analysis An Integrative Approach 1st Edition Sanders Test BankHeatherAllencejrx100% (16)
- Honeycutt 2009 Chap 76Document6 pagesHoneycutt 2009 Chap 76Michelle Alejandra BrionesNo ratings yet
- Gattaca PavelDocument3 pagesGattaca PavelpavelofficialNo ratings yet
- Increased Production of Azadirachtin From An Improved Method of Androgenic Cultures of A Medicinal Tree Azadirachta Indica A. JussDocument9 pagesIncreased Production of Azadirachtin From An Improved Method of Androgenic Cultures of A Medicinal Tree Azadirachta Indica A. JussPrathamesh BolajNo ratings yet
- Post Feb Mock Plan: Topics 1-4 PAPER 1 Assessment Focus RPDocument61 pagesPost Feb Mock Plan: Topics 1-4 PAPER 1 Assessment Focus RPEllisNo ratings yet
- Coconut Genetic ResourcesDocument797 pagesCoconut Genetic ResourcesJohnDavidNo ratings yet
- sc1c02627 Si 001Document16 pagessc1c02627 Si 001Maii MohammedNo ratings yet
- Adapterama Iii: Quadruple-Indexed, Triple-Enzyme Radseq Libraries For About $1usd Per Sample (3rad)Document35 pagesAdapterama Iii: Quadruple-Indexed, Triple-Enzyme Radseq Libraries For About $1usd Per Sample (3rad)OanaNo ratings yet
- Development TasksDocument4 pagesDevelopment TasksgretutegeruteNo ratings yet