
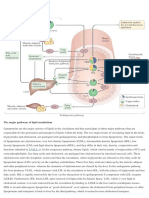
Advances in HDL: Much More Than Lipid Transporters: Molecular Sciences
Advances in HDL: Much More Than Lipid Transporters: Molecular Sciences
You might also like
- Books About Nootropics. Arushanian Final With The CoverDocument113 pagesBooks About Nootropics. Arushanian Final With The CoverAudreyNo ratings yet
- Hospice Care (End of Life Care) For Terminal Illness - Case Study SampleDocument19 pagesHospice Care (End of Life Care) For Terminal Illness - Case Study Samplepeng kulongNo ratings yet
- Statins Stimulate Atherosclerosis and Heart Failure Pharmacological MechanismsDocument12 pagesStatins Stimulate Atherosclerosis and Heart Failure Pharmacological MechanismsSonata DaniatiekNo ratings yet
- A Safe, Natural Way To End AddictionDocument9 pagesA Safe, Natural Way To End AddictionnupathwaysNo ratings yet
- HDL in CKD-ThesisDocument27 pagesHDL in CKD-ThesisEndy SusenoNo ratings yet
- Reverse Cholesterol TransportDocument29 pagesReverse Cholesterol TransportSolomon RotimiNo ratings yet
- Meningkatkan Kolesterol HDL Paradigma Baru Dalam Pencegahan Penyakit KardiovaskularDocument10 pagesMeningkatkan Kolesterol HDL Paradigma Baru Dalam Pencegahan Penyakit KardiovaskularMega RatnasariNo ratings yet
- International Journal of Health Sciences and ResearchDocument8 pagesInternational Journal of Health Sciences and ResearchwakhidrizkaNo ratings yet
- Introduction To Lipids and LipoproteinsDocument24 pagesIntroduction To Lipids and LipoproteinsYoungFanjiensNo ratings yet
- Farmakoterapi DislipidemiaDocument52 pagesFarmakoterapi DislipidemialisaNo ratings yet
- Jurnal Inter 3Document15 pagesJurnal Inter 3Annisa YusrilianiNo ratings yet
- Bpjv07i02p399 409Document12 pagesBpjv07i02p399 409PNo ratings yet
- Disorders of Lipid Metabolism Full Article Tables FiguresDocument32 pagesDisorders of Lipid Metabolism Full Article Tables Figureshasnawati100% (1)
- 4.4.3 (Bioc) Lipoproteins: Chylomicrons, VLDL, LDL, and HDL: Biochemistry, TwoDocument22 pages4.4.3 (Bioc) Lipoproteins: Chylomicrons, VLDL, LDL, and HDL: Biochemistry, TwoAyro Business CenterNo ratings yet
- Abstract and IntroductionDocument14 pagesAbstract and Introductiondavid_lg6179No ratings yet
- Toward A Higher Standard: Raising HDL in Clinical PracticeDocument11 pagesToward A Higher Standard: Raising HDL in Clinical PracticeAlexandre Sant'AnaNo ratings yet
- Review: S. E. Borggreve, R. de Vries and R. P. F. DullaartDocument19 pagesReview: S. E. Borggreve, R. de Vries and R. P. F. DullaartFCiênciasNo ratings yet
- Jurnal 5Document5 pagesJurnal 5zahriz zaitunNo ratings yet
- Bandeali-Farmer2012 Article High-DensityLipoproteinAndAtheDocument7 pagesBandeali-Farmer2012 Article High-DensityLipoproteinAndAtheDainius JakubauskasNo ratings yet
- Pharmacology :MIT:Lees:8Document8 pagesPharmacology :MIT:Lees:8Louis FortunatoNo ratings yet
- Lipoproteins: SummaryDocument3 pagesLipoproteins: SummaryEzekiel Morena RiveraNo ratings yet
- Cardiology Update:: LDL Reduction and HDL Management 2012Document8 pagesCardiology Update:: LDL Reduction and HDL Management 2012senaringNo ratings yet
- High-Density Lipoproteins As Homeostatic Nanoparticles of Blood PlasmaDocument18 pagesHigh-Density Lipoproteins As Homeostatic Nanoparticles of Blood PlasmaBBD BBDNo ratings yet
- Biokimia LipidDocument6 pagesBiokimia Lipiddimensi warnaNo ratings yet
- Lipoprotein Metabolism Note-1Document8 pagesLipoprotein Metabolism Note-1TimilehinNo ratings yet
- Lifestyle Factors and Pharmacological Agents That Manage Hyperlipidemia Through Activation of Peroxisome Proliferator Activated ReceptorsDocument16 pagesLifestyle Factors and Pharmacological Agents That Manage Hyperlipidemia Through Activation of Peroxisome Proliferator Activated ReceptorsjebacNo ratings yet
- J. Lipid Res. 1984 Eisenberg 1017 58Document42 pagesJ. Lipid Res. 1984 Eisenberg 1017 58RainolNo ratings yet
- Biochemistry of Lipoprotein & DyslipidemiaDocument79 pagesBiochemistry of Lipoprotein & DyslipidemiaDamal An NasherNo ratings yet
- Caso Clinico 30Document8 pagesCaso Clinico 30Ariel SanNo ratings yet
- Poster-2-Makalah Ilmiah Glycine Soja+glycine Max1-EditDocument7 pagesPoster-2-Makalah Ilmiah Glycine Soja+glycine Max1-EditArief SurizaNo ratings yet
- Biochem 9-10Document2 pagesBiochem 9-10Christine Arellano IIINo ratings yet
- Lipids Lipoproteins ApolipoproteinsDocument26 pagesLipids Lipoproteins ApolipoproteinssakuraleeshaoranNo ratings yet
- Lipoprotein MetabolismDocument23 pagesLipoprotein MetabolismDarien LiewNo ratings yet
- Biomarker 3Document7 pagesBiomarker 3Raluca Andreea AxinteNo ratings yet
- Nanotechnology For Synthetic High-Density LipoproteinsDocument8 pagesNanotechnology For Synthetic High-Density Lipoproteinsdev_sNo ratings yet
- Lipoprotein Group 5Document46 pagesLipoprotein Group 5SharNo ratings yet
- What Are LipoproteinsDocument18 pagesWhat Are Lipoproteinsarsal1cheema-88705No ratings yet
- Effects of Dietary Flavonoids On Reverse Cholesterol Transport, HDL Metabolism, and HDL FunctionDocument14 pagesEffects of Dietary Flavonoids On Reverse Cholesterol Transport, HDL Metabolism, and HDL FunctionYoungFanjiensNo ratings yet
- Biochemistry LipoproteinDocument84 pagesBiochemistry LipoproteiniqiqiqiqiqNo ratings yet
- Review Article The Major Pathways of Lipids (Triglyceride and Cholesterol) and Lipoprotein MetabolismDocument12 pagesReview Article The Major Pathways of Lipids (Triglyceride and Cholesterol) and Lipoprotein MetabolismRe: AlChemist BrotherhoodNo ratings yet
- Module 11 Lipids 1Document10 pagesModule 11 Lipids 1cassseeeyyyNo ratings yet
- Lipioprotein Lec11 PDFDocument44 pagesLipioprotein Lec11 PDFحسن محمد سعيد جاسمNo ratings yet
- چمستريDocument39 pagesچمستريn99xsbvjqkNo ratings yet
- كيمياءDocument7 pagesكيمياءn99xsbvjqkNo ratings yet
- Biochem1 Practical6Document8 pagesBiochem1 Practical6puhranee nathanNo ratings yet
- Figure - PMCDocument1 pageFigure - PMCHareesh ChanderNo ratings yet
- Figure - PMC 2Document1 pageFigure - PMC 2Hareesh ChanderNo ratings yet
- Complex Proteins - Glyco, Lipo, PhosphoDocument19 pagesComplex Proteins - Glyco, Lipo, Phosphodc8mypycmdNo ratings yet
- Digestion of Lipids. Transport Forms of LipidsDocument91 pagesDigestion of Lipids. Transport Forms of LipidsAhmed FtsNo ratings yet
- Review: - ACC RegulationDocument41 pagesReview: - ACC RegulationNico AturdidoNo ratings yet
- Metabolism of HDLDocument20 pagesMetabolism of HDLAisha HasanNo ratings yet
- Lipid Metabolism - 3rd LectureDocument38 pagesLipid Metabolism - 3rd LectureLe Uyen Nguyen100% (1)
- Lipid MetabolismDocument19 pagesLipid MetabolismDaniel PanjaitanNo ratings yet
- Lecture 11. Blood Biochemistry. Erythrocytes Metabolism. HemoglobinDocument27 pagesLecture 11. Blood Biochemistry. Erythrocytes Metabolism. HemoglobinВіталій Михайлович НечипорукNo ratings yet
- Annals of Clinical Biochemistry: International Journal of Laboratory MedicineDocument93 pagesAnnals of Clinical Biochemistry: International Journal of Laboratory MedicinePaula VillaNo ratings yet
- Metabolism of Lipids And: LipoproteinsDocument50 pagesMetabolism of Lipids And: LipoproteinsRichoco SapoetraNo ratings yet
- Energy Production 2. Energy Storage 3. Energy UtilizationDocument14 pagesEnergy Production 2. Energy Storage 3. Energy UtilizationLALITH SAI KNo ratings yet
- High-Density Lipoprotein Cholesterol - The Next Battle Front in The Fight Against Hear T DiseaseDocument8 pagesHigh-Density Lipoprotein Cholesterol - The Next Battle Front in The Fight Against Hear T DiseaseRainolNo ratings yet
- Lipoprotein - WikipediaDocument38 pagesLipoprotein - WikipediaTejaswiNo ratings yet
- Dyslipidemia PDFDocument28 pagesDyslipidemia PDFwilliaNo ratings yet
- Atherogenesis Accumulation of Lipoprotein-Derived Lipids Including Cholesteryl Ester (CE) andDocument6 pagesAtherogenesis Accumulation of Lipoprotein-Derived Lipids Including Cholesteryl Ester (CE) andrajeshmangalNo ratings yet
- Fast Facts: Hyperlipidemia: Bringing clarity to lipid managementFrom EverandFast Facts: Hyperlipidemia: Bringing clarity to lipid managementNo ratings yet
- High-Density Lipoproteins: Structure, Metabolism, Function and TherapeuticsFrom EverandHigh-Density Lipoproteins: Structure, Metabolism, Function and TherapeuticsNo ratings yet
- Diabetic Vascular Diseases: Molecular Mechanisms and Therapeutic StrategiesDocument29 pagesDiabetic Vascular Diseases: Molecular Mechanisms and Therapeutic StrategiesBBD BBDNo ratings yet
- Textbook of Vascular Medicine: Rhian M. Touyz Christian DellesDocument497 pagesTextbook of Vascular Medicine: Rhian M. Touyz Christian DellesBBD BBDNo ratings yet
- 1 s2.0 S2452302X20303946 MainDocument15 pages1 s2.0 S2452302X20303946 MainBBD BBDNo ratings yet
- Diseases 09 00066 v2Document19 pagesDiseases 09 00066 v2BBD BBDNo ratings yet
- Children 10 00695Document16 pagesChildren 10 00695BBD BBDNo ratings yet
- H PyloriDocument7 pagesH PyloriBBD BBDNo ratings yet
- H Pylori RGB Single Page AW2022Document16 pagesH Pylori RGB Single Page AW2022BBD BBDNo ratings yet
- Asthma - Types, Causes, Symptoms, Diagnosis & TreatmentDocument14 pagesAsthma - Types, Causes, Symptoms, Diagnosis & TreatmentBBD BBDNo ratings yet
- ResearchDocument21 pagesResearchBBD BBDNo ratings yet
- Advanced Drug Delivery ReviewsDocument26 pagesAdvanced Drug Delivery ReviewsBBD BBDNo ratings yet
- Clinical Nutrition ESPENDocument15 pagesClinical Nutrition ESPENBBD BBDNo ratings yet
- NutrientsDocument12 pagesNutrientsBBD BBDNo ratings yet
- 1 s2.0 S1043661823001366 MainDocument12 pages1 s2.0 S1043661823001366 MainBBD BBDNo ratings yet
- An Update On Asthma DiagnosisDocument8 pagesAn Update On Asthma DiagnosisBBD BBDNo ratings yet
- Comment: See Page E218Document2 pagesComment: See Page E218BBD BBDNo ratings yet
- AntibioticsDocument14 pagesAntibioticsBBD BBDNo ratings yet
- Analysis of Antibiotic Consumption by Aware Classi Fication in Shandong Province, China, 2012 - 2019: A Panel Data AnalysisDocument8 pagesAnalysis of Antibiotic Consumption by Aware Classi Fication in Shandong Province, China, 2012 - 2019: A Panel Data AnalysisBBD BBDNo ratings yet
- The Changing Landscape of Children's Diet and Nutrition: New Threats, New OpportunitiesDocument11 pagesThe Changing Landscape of Children's Diet and Nutrition: New Threats, New OpportunitiesBBD BBDNo ratings yet
- BL 39Document3 pagesBL 39YAMINIPRIYANNo ratings yet
- Stability Indicating RP-HPLC Method For Simultaneous Determination of Perindopril and Indapamide in Pharmaceutical Dosage FormDocument9 pagesStability Indicating RP-HPLC Method For Simultaneous Determination of Perindopril and Indapamide in Pharmaceutical Dosage FormBoovizhikannan ThangabalanNo ratings yet
- Psychiatric Practice TestDocument4 pagesPsychiatric Practice TestARIS100% (2)
- BMC Psychiatry: Arne E Vaaler, Gunnar Morken, Olav M Linaker, Trond Sand, Kjell A Kvistad and Geir BråthenDocument6 pagesBMC Psychiatry: Arne E Vaaler, Gunnar Morken, Olav M Linaker, Trond Sand, Kjell A Kvistad and Geir Bråthenaria tristayanthiNo ratings yet
- Schizophrenia in Adults - Epidemiology and Pathogenesis - UpToDateDocument23 pagesSchizophrenia in Adults - Epidemiology and Pathogenesis - UpToDateJorge AlvizoNo ratings yet
- Local Anesthesia Manual: Loma Linda University School of DentistryDocument81 pagesLocal Anesthesia Manual: Loma Linda University School of DentistryRichiene Bartolome CustorioNo ratings yet
- Stephan Heckers Catatonia 2023Document6 pagesStephan Heckers Catatonia 2023kendoca23No ratings yet
- Chapter 6 - Endocrine System 2Document12 pagesChapter 6 - Endocrine System 2AyeshaNo ratings yet
- Pharmaceutics-Ii (2131) Theory: 75 Hours (3 Hours/week), Practical: 75 Hours (Total 6 Hours/Week, 3 Hours/Week Per Batch)Document21 pagesPharmaceutics-Ii (2131) Theory: 75 Hours (3 Hours/week), Practical: 75 Hours (Total 6 Hours/Week, 3 Hours/Week Per Batch)Harshal DodkeNo ratings yet
- Fact UrasDocument6 pagesFact UrasRonald Salazar CuzcanoNo ratings yet
- PharmacyDocument4 pagesPharmacyEduCartNo ratings yet
- Providing Comfort During LaborDocument11 pagesProviding Comfort During LaborAndrea Marie SevillaNo ratings yet
- Handbook of Wildlife Chemical Immobilization, 5th EditionDocument596 pagesHandbook of Wildlife Chemical Immobilization, 5th EditionDiego RiosNo ratings yet
- Thank You Truist. Smart Fund's Response To Truist Securities' Defence of IGMS Bear ThesisDocument21 pagesThank You Truist. Smart Fund's Response To Truist Securities' Defence of IGMS Bear Thesisjulia skripka-serryNo ratings yet
- Disorders of Autonomic Nervous System: Chair PersonsDocument37 pagesDisorders of Autonomic Nervous System: Chair PersonswarunkumarNo ratings yet
- Severe Dopaminergic Neurotoxicity in Primates After A Common Recreational Dose Regimen of MDMA ('Ecstasy')Document5 pagesSevere Dopaminergic Neurotoxicity in Primates After A Common Recreational Dose Regimen of MDMA ('Ecstasy')asdadasdasNo ratings yet
- Sleep and WakefulnessDocument12 pagesSleep and WakefulnessJill MehtaNo ratings yet
- History of Hematology (To Be Edited)Document6 pagesHistory of Hematology (To Be Edited)Reca Marie FRIASNo ratings yet
- E-Cigarettes A Rising Trend Among YouthDocument8 pagesE-Cigarettes A Rising Trend Among YouthAsd EfgNo ratings yet
- Ho Notes Part 1 - Medical Emergencies Notes 2.0Document7 pagesHo Notes Part 1 - Medical Emergencies Notes 2.0Amin MasromNo ratings yet
- Amputation of Tail in Buff JournalDocument9 pagesAmputation of Tail in Buff JournalKhawaja usmanNo ratings yet
- The Control of LaborDocument7 pagesThe Control of LaborDR RISKA WAHYUNo ratings yet
- Charm Pour Detection Residu Ds Lait PDFDocument2 pagesCharm Pour Detection Residu Ds Lait PDFhassanNo ratings yet
- All DrugsDocument150 pagesAll Drugsপ্রান্ত সরকারNo ratings yet
- Electrolytes PDFDocument5 pagesElectrolytes PDFFrances FranciscoNo ratings yet
- The Open Anesthesia JournalDocument11 pagesThe Open Anesthesia JournalmikoNo ratings yet
- Procedural Sedation RecordDocument2 pagesProcedural Sedation RecordRosanne AguilarNo ratings yet










































































































Download as pdf or txt
You might also like
- Books About Nootropics. Arushanian Final With The CoverDocument113 pagesBooks About Nootropics. Arushanian Final With The CoverAudreyNo ratings yet
- Hospice Care (End of Life Care) For Terminal Illness - Case Study SampleDocument19 pagesHospice Care (End of Life Care) For Terminal Illness - Case Study Samplepeng kulongNo ratings yet
- Statins Stimulate Atherosclerosis and Heart Failure Pharmacological MechanismsDocument12 pagesStatins Stimulate Atherosclerosis and Heart Failure Pharmacological MechanismsSonata DaniatiekNo ratings yet
- A Safe, Natural Way To End AddictionDocument9 pagesA Safe, Natural Way To End AddictionnupathwaysNo ratings yet
- HDL in CKD-ThesisDocument27 pagesHDL in CKD-ThesisEndy SusenoNo ratings yet
- Reverse Cholesterol TransportDocument29 pagesReverse Cholesterol TransportSolomon RotimiNo ratings yet
- Meningkatkan Kolesterol HDL Paradigma Baru Dalam Pencegahan Penyakit KardiovaskularDocument10 pagesMeningkatkan Kolesterol HDL Paradigma Baru Dalam Pencegahan Penyakit KardiovaskularMega RatnasariNo ratings yet
- International Journal of Health Sciences and ResearchDocument8 pagesInternational Journal of Health Sciences and ResearchwakhidrizkaNo ratings yet
- Introduction To Lipids and LipoproteinsDocument24 pagesIntroduction To Lipids and LipoproteinsYoungFanjiensNo ratings yet
- Farmakoterapi DislipidemiaDocument52 pagesFarmakoterapi DislipidemialisaNo ratings yet
- Jurnal Inter 3Document15 pagesJurnal Inter 3Annisa YusrilianiNo ratings yet
- Bpjv07i02p399 409Document12 pagesBpjv07i02p399 409PNo ratings yet
- Disorders of Lipid Metabolism Full Article Tables FiguresDocument32 pagesDisorders of Lipid Metabolism Full Article Tables Figureshasnawati100% (1)
- 4.4.3 (Bioc) Lipoproteins: Chylomicrons, VLDL, LDL, and HDL: Biochemistry, TwoDocument22 pages4.4.3 (Bioc) Lipoproteins: Chylomicrons, VLDL, LDL, and HDL: Biochemistry, TwoAyro Business CenterNo ratings yet
- Abstract and IntroductionDocument14 pagesAbstract and Introductiondavid_lg6179No ratings yet
- Toward A Higher Standard: Raising HDL in Clinical PracticeDocument11 pagesToward A Higher Standard: Raising HDL in Clinical PracticeAlexandre Sant'AnaNo ratings yet
- Review: S. E. Borggreve, R. de Vries and R. P. F. DullaartDocument19 pagesReview: S. E. Borggreve, R. de Vries and R. P. F. DullaartFCiênciasNo ratings yet
- Jurnal 5Document5 pagesJurnal 5zahriz zaitunNo ratings yet
- Bandeali-Farmer2012 Article High-DensityLipoproteinAndAtheDocument7 pagesBandeali-Farmer2012 Article High-DensityLipoproteinAndAtheDainius JakubauskasNo ratings yet
- Pharmacology :MIT:Lees:8Document8 pagesPharmacology :MIT:Lees:8Louis FortunatoNo ratings yet
- Lipoproteins: SummaryDocument3 pagesLipoproteins: SummaryEzekiel Morena RiveraNo ratings yet
- Cardiology Update:: LDL Reduction and HDL Management 2012Document8 pagesCardiology Update:: LDL Reduction and HDL Management 2012senaringNo ratings yet
- High-Density Lipoproteins As Homeostatic Nanoparticles of Blood PlasmaDocument18 pagesHigh-Density Lipoproteins As Homeostatic Nanoparticles of Blood PlasmaBBD BBDNo ratings yet
- Biokimia LipidDocument6 pagesBiokimia Lipiddimensi warnaNo ratings yet
- Lipoprotein Metabolism Note-1Document8 pagesLipoprotein Metabolism Note-1TimilehinNo ratings yet
- Lifestyle Factors and Pharmacological Agents That Manage Hyperlipidemia Through Activation of Peroxisome Proliferator Activated ReceptorsDocument16 pagesLifestyle Factors and Pharmacological Agents That Manage Hyperlipidemia Through Activation of Peroxisome Proliferator Activated ReceptorsjebacNo ratings yet
- J. Lipid Res. 1984 Eisenberg 1017 58Document42 pagesJ. Lipid Res. 1984 Eisenberg 1017 58RainolNo ratings yet
- Biochemistry of Lipoprotein & DyslipidemiaDocument79 pagesBiochemistry of Lipoprotein & DyslipidemiaDamal An NasherNo ratings yet
- Caso Clinico 30Document8 pagesCaso Clinico 30Ariel SanNo ratings yet
- Poster-2-Makalah Ilmiah Glycine Soja+glycine Max1-EditDocument7 pagesPoster-2-Makalah Ilmiah Glycine Soja+glycine Max1-EditArief SurizaNo ratings yet
- Biochem 9-10Document2 pagesBiochem 9-10Christine Arellano IIINo ratings yet
- Lipids Lipoproteins ApolipoproteinsDocument26 pagesLipids Lipoproteins ApolipoproteinssakuraleeshaoranNo ratings yet
- Lipoprotein MetabolismDocument23 pagesLipoprotein MetabolismDarien LiewNo ratings yet
- Biomarker 3Document7 pagesBiomarker 3Raluca Andreea AxinteNo ratings yet
- Nanotechnology For Synthetic High-Density LipoproteinsDocument8 pagesNanotechnology For Synthetic High-Density Lipoproteinsdev_sNo ratings yet
- Lipoprotein Group 5Document46 pagesLipoprotein Group 5SharNo ratings yet
- What Are LipoproteinsDocument18 pagesWhat Are Lipoproteinsarsal1cheema-88705No ratings yet
- Effects of Dietary Flavonoids On Reverse Cholesterol Transport, HDL Metabolism, and HDL FunctionDocument14 pagesEffects of Dietary Flavonoids On Reverse Cholesterol Transport, HDL Metabolism, and HDL FunctionYoungFanjiensNo ratings yet
- Biochemistry LipoproteinDocument84 pagesBiochemistry LipoproteiniqiqiqiqiqNo ratings yet
- Review Article The Major Pathways of Lipids (Triglyceride and Cholesterol) and Lipoprotein MetabolismDocument12 pagesReview Article The Major Pathways of Lipids (Triglyceride and Cholesterol) and Lipoprotein MetabolismRe: AlChemist BrotherhoodNo ratings yet
- Module 11 Lipids 1Document10 pagesModule 11 Lipids 1cassseeeyyyNo ratings yet
- Lipioprotein Lec11 PDFDocument44 pagesLipioprotein Lec11 PDFحسن محمد سعيد جاسمNo ratings yet
- چمستريDocument39 pagesچمستريn99xsbvjqkNo ratings yet
- كيمياءDocument7 pagesكيمياءn99xsbvjqkNo ratings yet
- Biochem1 Practical6Document8 pagesBiochem1 Practical6puhranee nathanNo ratings yet
- Figure - PMCDocument1 pageFigure - PMCHareesh ChanderNo ratings yet
- Figure - PMC 2Document1 pageFigure - PMC 2Hareesh ChanderNo ratings yet
- Complex Proteins - Glyco, Lipo, PhosphoDocument19 pagesComplex Proteins - Glyco, Lipo, Phosphodc8mypycmdNo ratings yet
- Digestion of Lipids. Transport Forms of LipidsDocument91 pagesDigestion of Lipids. Transport Forms of LipidsAhmed FtsNo ratings yet
- Review: - ACC RegulationDocument41 pagesReview: - ACC RegulationNico AturdidoNo ratings yet
- Metabolism of HDLDocument20 pagesMetabolism of HDLAisha HasanNo ratings yet
- Lipid Metabolism - 3rd LectureDocument38 pagesLipid Metabolism - 3rd LectureLe Uyen Nguyen100% (1)
- Lipid MetabolismDocument19 pagesLipid MetabolismDaniel PanjaitanNo ratings yet
- Lecture 11. Blood Biochemistry. Erythrocytes Metabolism. HemoglobinDocument27 pagesLecture 11. Blood Biochemistry. Erythrocytes Metabolism. HemoglobinВіталій Михайлович НечипорукNo ratings yet
- Annals of Clinical Biochemistry: International Journal of Laboratory MedicineDocument93 pagesAnnals of Clinical Biochemistry: International Journal of Laboratory MedicinePaula VillaNo ratings yet
- Metabolism of Lipids And: LipoproteinsDocument50 pagesMetabolism of Lipids And: LipoproteinsRichoco SapoetraNo ratings yet
- Energy Production 2. Energy Storage 3. Energy UtilizationDocument14 pagesEnergy Production 2. Energy Storage 3. Energy UtilizationLALITH SAI KNo ratings yet
- High-Density Lipoprotein Cholesterol - The Next Battle Front in The Fight Against Hear T DiseaseDocument8 pagesHigh-Density Lipoprotein Cholesterol - The Next Battle Front in The Fight Against Hear T DiseaseRainolNo ratings yet
- Lipoprotein - WikipediaDocument38 pagesLipoprotein - WikipediaTejaswiNo ratings yet
- Dyslipidemia PDFDocument28 pagesDyslipidemia PDFwilliaNo ratings yet
- Atherogenesis Accumulation of Lipoprotein-Derived Lipids Including Cholesteryl Ester (CE) andDocument6 pagesAtherogenesis Accumulation of Lipoprotein-Derived Lipids Including Cholesteryl Ester (CE) andrajeshmangalNo ratings yet
- Fast Facts: Hyperlipidemia: Bringing clarity to lipid managementFrom EverandFast Facts: Hyperlipidemia: Bringing clarity to lipid managementNo ratings yet
- High-Density Lipoproteins: Structure, Metabolism, Function and TherapeuticsFrom EverandHigh-Density Lipoproteins: Structure, Metabolism, Function and TherapeuticsNo ratings yet
- Diabetic Vascular Diseases: Molecular Mechanisms and Therapeutic StrategiesDocument29 pagesDiabetic Vascular Diseases: Molecular Mechanisms and Therapeutic StrategiesBBD BBDNo ratings yet
- Textbook of Vascular Medicine: Rhian M. Touyz Christian DellesDocument497 pagesTextbook of Vascular Medicine: Rhian M. Touyz Christian DellesBBD BBDNo ratings yet
- 1 s2.0 S2452302X20303946 MainDocument15 pages1 s2.0 S2452302X20303946 MainBBD BBDNo ratings yet
- Diseases 09 00066 v2Document19 pagesDiseases 09 00066 v2BBD BBDNo ratings yet
- Children 10 00695Document16 pagesChildren 10 00695BBD BBDNo ratings yet
- H PyloriDocument7 pagesH PyloriBBD BBDNo ratings yet
- H Pylori RGB Single Page AW2022Document16 pagesH Pylori RGB Single Page AW2022BBD BBDNo ratings yet
- Asthma - Types, Causes, Symptoms, Diagnosis & TreatmentDocument14 pagesAsthma - Types, Causes, Symptoms, Diagnosis & TreatmentBBD BBDNo ratings yet
- ResearchDocument21 pagesResearchBBD BBDNo ratings yet
- Advanced Drug Delivery ReviewsDocument26 pagesAdvanced Drug Delivery ReviewsBBD BBDNo ratings yet
- Clinical Nutrition ESPENDocument15 pagesClinical Nutrition ESPENBBD BBDNo ratings yet
- NutrientsDocument12 pagesNutrientsBBD BBDNo ratings yet
- 1 s2.0 S1043661823001366 MainDocument12 pages1 s2.0 S1043661823001366 MainBBD BBDNo ratings yet
- An Update On Asthma DiagnosisDocument8 pagesAn Update On Asthma DiagnosisBBD BBDNo ratings yet
- Comment: See Page E218Document2 pagesComment: See Page E218BBD BBDNo ratings yet
- AntibioticsDocument14 pagesAntibioticsBBD BBDNo ratings yet
- Analysis of Antibiotic Consumption by Aware Classi Fication in Shandong Province, China, 2012 - 2019: A Panel Data AnalysisDocument8 pagesAnalysis of Antibiotic Consumption by Aware Classi Fication in Shandong Province, China, 2012 - 2019: A Panel Data AnalysisBBD BBDNo ratings yet
- The Changing Landscape of Children's Diet and Nutrition: New Threats, New OpportunitiesDocument11 pagesThe Changing Landscape of Children's Diet and Nutrition: New Threats, New OpportunitiesBBD BBDNo ratings yet
- BL 39Document3 pagesBL 39YAMINIPRIYANNo ratings yet
- Stability Indicating RP-HPLC Method For Simultaneous Determination of Perindopril and Indapamide in Pharmaceutical Dosage FormDocument9 pagesStability Indicating RP-HPLC Method For Simultaneous Determination of Perindopril and Indapamide in Pharmaceutical Dosage FormBoovizhikannan ThangabalanNo ratings yet
- Psychiatric Practice TestDocument4 pagesPsychiatric Practice TestARIS100% (2)
- BMC Psychiatry: Arne E Vaaler, Gunnar Morken, Olav M Linaker, Trond Sand, Kjell A Kvistad and Geir BråthenDocument6 pagesBMC Psychiatry: Arne E Vaaler, Gunnar Morken, Olav M Linaker, Trond Sand, Kjell A Kvistad and Geir Bråthenaria tristayanthiNo ratings yet
- Schizophrenia in Adults - Epidemiology and Pathogenesis - UpToDateDocument23 pagesSchizophrenia in Adults - Epidemiology and Pathogenesis - UpToDateJorge AlvizoNo ratings yet
- Local Anesthesia Manual: Loma Linda University School of DentistryDocument81 pagesLocal Anesthesia Manual: Loma Linda University School of DentistryRichiene Bartolome CustorioNo ratings yet
- Stephan Heckers Catatonia 2023Document6 pagesStephan Heckers Catatonia 2023kendoca23No ratings yet
- Chapter 6 - Endocrine System 2Document12 pagesChapter 6 - Endocrine System 2AyeshaNo ratings yet
- Pharmaceutics-Ii (2131) Theory: 75 Hours (3 Hours/week), Practical: 75 Hours (Total 6 Hours/Week, 3 Hours/Week Per Batch)Document21 pagesPharmaceutics-Ii (2131) Theory: 75 Hours (3 Hours/week), Practical: 75 Hours (Total 6 Hours/Week, 3 Hours/Week Per Batch)Harshal DodkeNo ratings yet
- Fact UrasDocument6 pagesFact UrasRonald Salazar CuzcanoNo ratings yet
- PharmacyDocument4 pagesPharmacyEduCartNo ratings yet
- Providing Comfort During LaborDocument11 pagesProviding Comfort During LaborAndrea Marie SevillaNo ratings yet
- Handbook of Wildlife Chemical Immobilization, 5th EditionDocument596 pagesHandbook of Wildlife Chemical Immobilization, 5th EditionDiego RiosNo ratings yet
- Thank You Truist. Smart Fund's Response To Truist Securities' Defence of IGMS Bear ThesisDocument21 pagesThank You Truist. Smart Fund's Response To Truist Securities' Defence of IGMS Bear Thesisjulia skripka-serryNo ratings yet
- Disorders of Autonomic Nervous System: Chair PersonsDocument37 pagesDisorders of Autonomic Nervous System: Chair PersonswarunkumarNo ratings yet
- Severe Dopaminergic Neurotoxicity in Primates After A Common Recreational Dose Regimen of MDMA ('Ecstasy')Document5 pagesSevere Dopaminergic Neurotoxicity in Primates After A Common Recreational Dose Regimen of MDMA ('Ecstasy')asdadasdasNo ratings yet
- Sleep and WakefulnessDocument12 pagesSleep and WakefulnessJill MehtaNo ratings yet
- History of Hematology (To Be Edited)Document6 pagesHistory of Hematology (To Be Edited)Reca Marie FRIASNo ratings yet
- E-Cigarettes A Rising Trend Among YouthDocument8 pagesE-Cigarettes A Rising Trend Among YouthAsd EfgNo ratings yet
- Ho Notes Part 1 - Medical Emergencies Notes 2.0Document7 pagesHo Notes Part 1 - Medical Emergencies Notes 2.0Amin MasromNo ratings yet
- Amputation of Tail in Buff JournalDocument9 pagesAmputation of Tail in Buff JournalKhawaja usmanNo ratings yet
- The Control of LaborDocument7 pagesThe Control of LaborDR RISKA WAHYUNo ratings yet
- Charm Pour Detection Residu Ds Lait PDFDocument2 pagesCharm Pour Detection Residu Ds Lait PDFhassanNo ratings yet
- All DrugsDocument150 pagesAll Drugsপ্রান্ত সরকারNo ratings yet
- Electrolytes PDFDocument5 pagesElectrolytes PDFFrances FranciscoNo ratings yet
- The Open Anesthesia JournalDocument11 pagesThe Open Anesthesia JournalmikoNo ratings yet
- Procedural Sedation RecordDocument2 pagesProcedural Sedation RecordRosanne AguilarNo ratings yet