Download as pdf or txt
You might also like
- Fast Facts: Diagnosing Amyotrophic Lateral Sclerosis: Clinical wisdom to facilitate faster diagnosisFrom EverandFast Facts: Diagnosing Amyotrophic Lateral Sclerosis: Clinical wisdom to facilitate faster diagnosisNo ratings yet
- Case 40-2021 - A 9-Year-Old - Enf - MoyamoyaDocument10 pagesCase 40-2021 - A 9-Year-Old - Enf - MoyamoyaJuan C. Salazar PajaresNo ratings yet
- Loss of The N-Myc Oncogene in A Patient With A Small Interstitial Deletion of The Short Arm of ChromosomeDocument5 pagesLoss of The N-Myc Oncogene in A Patient With A Small Interstitial Deletion of The Short Arm of ChromosomeantonioopNo ratings yet
- Algoritmo Diferencial de Debilidad Proximal en Paciente Con Miopatia Adquirida Por Nemalina.Document7 pagesAlgoritmo Diferencial de Debilidad Proximal en Paciente Con Miopatia Adquirida Por Nemalina.Farid Santiago Abedrabbo LombeydaNo ratings yet
- Kleine-Levin Syndrome-A Systematic Study of 108 PatientsDocument12 pagesKleine-Levin Syndrome-A Systematic Study of 108 PatientsChon ChiNo ratings yet
- An Example of The Utility of Genomic Analysis For Fast and Accurate Clinical Diagnosis of Complex Rare PhenotypesDocument8 pagesAn Example of The Utility of Genomic Analysis For Fast and Accurate Clinical Diagnosis of Complex Rare PhenotypesGlauce L TrevisanNo ratings yet
- Psychogenic Non-Epileptic Seizures: Indian Pediatrics November 2004Document4 pagesPsychogenic Non-Epileptic Seizures: Indian Pediatrics November 2004Nico NicoNo ratings yet
- Kolicheski Et Al-2017-Journal of Veterinary Internal MedicineDocument9 pagesKolicheski Et Al-2017-Journal of Veterinary Internal MedicineDulce Lucia ChacónNo ratings yet
- Essential Tremor: Clinical PracticeDocument9 pagesEssential Tremor: Clinical PracticeDaniel Torres100% (1)
- Wilson's Disease - The Problem of DelayedDocument5 pagesWilson's Disease - The Problem of DelayedBarry SeeboNo ratings yet
- Cervical DyatoniaDocument6 pagesCervical DyatoniaEmad Moody KaremNo ratings yet
- Five New Cases and Review of Nine Previously Published CasesDocument8 pagesFive New Cases and Review of Nine Previously Published CasesBogdan SoptereanNo ratings yet
- Screenshot 2023-09-17 at 3.26.16 PMDocument10 pagesScreenshot 2023-09-17 at 3.26.16 PMThana SnNo ratings yet
- Marinesco-Sjo Gren Syndrome - 2013Document5 pagesMarinesco-Sjo Gren Syndrome - 2013Irfan RazaNo ratings yet
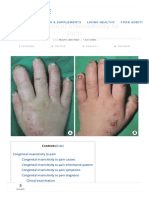
- Congenital Insensitivity To Pain-A Case Report With Review of LiteratureDocument3 pagesCongenital Insensitivity To Pain-A Case Report With Review of LiteratureJenirose C. GayagayNo ratings yet
- ResumenDocument5 pagesResumenPaulette BuñayNo ratings yet
- Estudio Clínico-Neurofisiológico de 70 Enfermos Con Ataxia Espinocerebelosa Tipo 2Document7 pagesEstudio Clínico-Neurofisiológico de 70 Enfermos Con Ataxia Espinocerebelosa Tipo 2David Fernando Ardila MoraNo ratings yet
- Letter To The Editor: Midline Field Defects and Hirschsprung DiseaseDocument2 pagesLetter To The Editor: Midline Field Defects and Hirschsprung DiseaseAraNo ratings yet
- 2009 BowsherDocument12 pages2009 BowsherMestrado EducacaoNo ratings yet
- Congenital Analgesia-Random NotesDocument5 pagesCongenital Analgesia-Random Notesltrathie3087No ratings yet
- Gratification Disorder ( Infantile Masturbation'') : A ReviewDocument2 pagesGratification Disorder ( Infantile Masturbation'') : A ReviewGibran UsamahNo ratings yet
- Clinicopathological Case Conference:: A 17 Year Old With Progressive Gait Difficulty and A Complex Family HistoryDocument3 pagesClinicopathological Case Conference:: A 17 Year Old With Progressive Gait Difficulty and A Complex Family HistoryChoi SpeaksNo ratings yet
- Harrower 1951Document14 pagesHarrower 1951Flavia ChNo ratings yet
- Journal RombergDocument7 pagesJournal RombergAnonymousssNo ratings yet
- Anaet 2Document3 pagesAnaet 2hidrisdmnk_747467972No ratings yet
- Carpe Diem Ruiz Protcol Grand Rounds 4Document10 pagesCarpe Diem Ruiz Protcol Grand Rounds 4John Christopher RuizNo ratings yet
- Carpe Diem Ruiz Protcol Grand Rounds 4Document9 pagesCarpe Diem Ruiz Protcol Grand Rounds 4John Christopher RuizNo ratings yet
- Fetal Trisomy 5 Mosaicism: Case Report and Literature ReviewDocument4 pagesFetal Trisomy 5 Mosaicism: Case Report and Literature ReviewjhscottNo ratings yet
- (Nigel G. Laing) The Sarcomere and Skeletal Muscle PDFDocument246 pages(Nigel G. Laing) The Sarcomere and Skeletal Muscle PDFVladimir Pablo Morales QuevedoNo ratings yet
- TMP 7 AC0Document11 pagesTMP 7 AC0FrontiersNo ratings yet
- Violencia Familiar y El Síndrome de Kluver-Bucy SMJ 1987Document5 pagesViolencia Familiar y El Síndrome de Kluver-Bucy SMJ 1987Jose MuñozNo ratings yet
- Hirschsprung's DiseaseDocument18 pagesHirschsprung's DiseaseanisyahNo ratings yet
- Approach To Febrile SeizuresDocument7 pagesApproach To Febrile SeizuresJohn Eric CudiaNo ratings yet
- Clinical Cases in Pediatric Peripheral NeuropathyDocument23 pagesClinical Cases in Pediatric Peripheral NeuropathyMateen ShukriNo ratings yet
- Alpers' DiseaseDocument5 pagesAlpers' DiseaseDANY PAUL BABYNo ratings yet
- Tolosa Hunt Syndrome: ArticlesDocument4 pagesTolosa Hunt Syndrome: ArticlesAyu Sylvia Axx HpNo ratings yet
- Case Report: Coexistence of Ankylosing Spondylitis and Neurofibromatosis Type 1Document4 pagesCase Report: Coexistence of Ankylosing Spondylitis and Neurofibromatosis Type 1Lupita Mora LobacoNo ratings yet
- Slavo Tine K 1996Document6 pagesSlavo Tine K 1996jahfdfgsdjad asdhsajhajdkNo ratings yet
- Neurological Complications Associated With Spontaneously Occurring Feline Diabetes MellitusDocument13 pagesNeurological Complications Associated With Spontaneously Occurring Feline Diabetes MellitusA J U N ANo ratings yet
- 03 Sarah OConnorDocument3 pages03 Sarah OConnorMateen ShukriNo ratings yet
- Plomp 1998Document7 pagesPlomp 1998dad dzd adaNo ratings yet
- Afectiuni Congenitale Ale FelinelorDocument34 pagesAfectiuni Congenitale Ale FelinelorVeaceslavNo ratings yet
- 10 1111@j 1440-1754 1995 tb00793 X PDFDocument4 pages10 1111@j 1440-1754 1995 tb00793 X PDFNdamale NicholasNo ratings yet
- ARB and PregnancySSDocument2 pagesARB and PregnancySSakhmadNo ratings yet
- Tactile Percepcion in Adults With Autism - A Multidimensional Psychophysical Study - Cascio 2008Document11 pagesTactile Percepcion in Adults With Autism - A Multidimensional Psychophysical Study - Cascio 2008Yahoska TijerinoNo ratings yet
- Bowman 1986Document8 pagesBowman 1986LNo ratings yet
- BFF 3Document4 pagesBFF 3الدكتور محمد علي الحصينيNo ratings yet
- Awareness of The Functioning of One's Own Limbs Mediated by The Insular Cortex?Document5 pagesAwareness of The Functioning of One's Own Limbs Mediated by The Insular Cortex?Julian GorositoNo ratings yet
- A Longitudinal Study of Back Pain and RadiologicalDocument5 pagesA Longitudinal Study of Back Pain and RadiologicalSumanta KamilaNo ratings yet
- Abstract: X-Linked Dystonia-Parkinsonism (XDP), or LubagDocument7 pagesAbstract: X-Linked Dystonia-Parkinsonism (XDP), or LubagKaila AbeledaNo ratings yet
- 425.full (1) OrthoDocument10 pages425.full (1) OrthoMA QureshiNo ratings yet
- Medina Et Al-2005-EpilepsiaDocument8 pagesMedina Et Al-2005-EpilepsiaGriselNo ratings yet
- Autosomal Dominant Microcephaly With Normal Intelligence, Short Palpebral Fissures, and Digital AnomaliesDocument6 pagesAutosomal Dominant Microcephaly With Normal Intelligence, Short Palpebral Fissures, and Digital Anomaliesjahfdfgsdjad asdhsajhajdkNo ratings yet
- Encefalitis LimbicaDocument11 pagesEncefalitis LimbicaRandy UlloaNo ratings yet
- Periodic Breathing in Patients With NALCN Mutations: Clinical ReportDocument4 pagesPeriodic Breathing in Patients With NALCN Mutations: Clinical ReportIvan VeriswanNo ratings yet
- Phenomenology and Classification of Dystonia - A Consensus Update (2013)Document11 pagesPhenomenology and Classification of Dystonia - A Consensus Update (2013)mysticmdNo ratings yet
- 1468-Article Text-3668-1-10-20170602Document8 pages1468-Article Text-3668-1-10-20170602ashighaNo ratings yet
- Greyson Alexander JNMD 2018Document4 pagesGreyson Alexander JNMD 2018Tibor AbaiNo ratings yet
- Morphometry of WS On ASDocument6 pagesMorphometry of WS On ASAdrien DuthéNo ratings yet
- Neurometabolic Hereditary Diseases of AdultsFrom EverandNeurometabolic Hereditary Diseases of AdultsAlessandro P. BurlinaNo ratings yet
- JNC 14939Document18 pagesJNC 14939wenshancheng2023No ratings yet
- 2004 Mutations in SCN9A, Encoding A Sodium Channel Alpha Subunit, in Patients With Primary ErythermalgiaDocument4 pages2004 Mutations in SCN9A, Encoding A Sodium Channel Alpha Subunit, in Patients With Primary Erythermalgiawenshancheng2023No ratings yet
- 2003 Sensory and electrophysiological properties of guinea‐pig sensory neurones expressing Nav 1.7 (PN1) Na+ channel α subunit proteinDocument12 pages2003 Sensory and electrophysiological properties of guinea‐pig sensory neurones expressing Nav 1.7 (PN1) Na+ channel α subunit proteinwenshancheng2023No ratings yet
- 1998 Slow Closed State Inactivation A Novel Mechanism UnderlyingDocument13 pages1998 Slow Closed State Inactivation A Novel Mechanism Underlyingwenshancheng2023No ratings yet
- 1997 Identification of PN1, A Predominant Voltage-Dependent Sodium Channel Expressed Principally in Peripheral NeuronsDocument6 pages1997 Identification of PN1, A Predominant Voltage-Dependent Sodium Channel Expressed Principally in Peripheral Neuronswenshancheng2023No ratings yet
- Chapter30 Electrical Kindling in Developing RatsDocument7 pagesChapter30 Electrical Kindling in Developing Ratswenshancheng2023No ratings yet
- Sodium Channel Blockers: A Patent Review (2010 - 2014)Document12 pagesSodium Channel Blockers: A Patent Review (2010 - 2014)BrandonMichaelWiseNo ratings yet
- Congenital Insensitivity To Pain Causes, Symptoms, Diagnosis & TDocument45 pagesCongenital Insensitivity To Pain Causes, Symptoms, Diagnosis & TPan DjeroNo ratings yet
- ChannelopathiesDocument18 pagesChannelopathiesJackyDanielsNo ratings yet
- Neuropathic Painn PDFDocument20 pagesNeuropathic Painn PDFkaktusNo ratings yet
- Neuropathic Pain: Emerging TreatmentsDocument11 pagesNeuropathic Pain: Emerging TreatmentsFarhatNo ratings yet
- ErythromelalgiaDocument4 pagesErythromelalgiaDr Reza ModarresNo ratings yet
- 1 Trigeminal Neuralgia An Overview From Pathophysiology To Pharmacological TreatmentDocument18 pages1 Trigeminal Neuralgia An Overview From Pathophysiology To Pharmacological TreatmentGabriela Elizabeth Gomez CardenasNo ratings yet
- Mechanical Allodynia: Invited ReviewDocument7 pagesMechanical Allodynia: Invited ReviewFabiano VargasNo ratings yet
- Lecture 1: Introducing The Power and Scope of Genetics Learning GoalsDocument8 pagesLecture 1: Introducing The Power and Scope of Genetics Learning GoalsAngelica SmithNo ratings yet
- Mutations of Voltage-Gated Sodium Channel Genes SCN1A and SCN2A in Epilepsy, Intellectual Disability, and AutismDocument19 pagesMutations of Voltage-Gated Sodium Channel Genes SCN1A and SCN2A in Epilepsy, Intellectual Disability, and Autismyasmin al ammariNo ratings yet
- Diagnosis and Management of Neuropathic ItchDocument12 pagesDiagnosis and Management of Neuropathic ItchTony Miguel Saba SabaNo ratings yet
- Voltage-Gated Ion Channels in The PNS: Novel Therapies For Neuropathic Pain?Document21 pagesVoltage-Gated Ion Channels in The PNS: Novel Therapies For Neuropathic Pain?Alicia Cerpa LunaNo ratings yet
- Pathophysiology of ParesthesiaDocument17 pagesPathophysiology of Paresthesiahardian0% (1)
- Nociceptor Sensory Neuron-Immune Interactions in Pain and InflammationDocument15 pagesNociceptor Sensory Neuron-Immune Interactions in Pain and InflammationRogerio MachadoNo ratings yet
- Jurnal Dr. PZDocument45 pagesJurnal Dr. PZRwsunbhio PratamaNo ratings yet
- Nav Structure-IntroductionDocument26 pagesNav Structure-IntroductionLorraine Joy dela CruzNo ratings yet
- Physiology of PainDocument30 pagesPhysiology of PainVaibhavJain100% (1)
- Pain Science in Practice - What Is Pain Neuroscience Part 1Document4 pagesPain Science in Practice - What Is Pain Neuroscience Part 1MathildeNo ratings yet
- Loza Huerta2021 Article Thallium SensitiveFluorescentADocument10 pagesLoza Huerta2021 Article Thallium SensitiveFluorescentAVivien JusztusNo ratings yet
- Mechanisms For Variable Expressivity of Inherited SCN1A Mutations Causing Dravet SyndromeDocument8 pagesMechanisms For Variable Expressivity of Inherited SCN1A Mutations Causing Dravet SyndromewoderickNo ratings yet
- Jospt 1Document3 pagesJospt 1Ian PeroniNo ratings yet
- SFN Expanding The Genetic Causes of Small Fiber Neuropathy SCN Genes and BeyondDocument13 pagesSFN Expanding The Genetic Causes of Small Fiber Neuropathy SCN Genes and BeyondOTTO VEGA VEGANo ratings yet