
Enriched Environment Ameliorates Dexamethasone
Enriched Environment Ameliorates Dexamethasone
You might also like
- Task Risk Assessment For RadiographyDocument5 pagesTask Risk Assessment For RadiographySanjeev Nair81% (21)
- Yamaha Owners Manual 40 50 HPDocument88 pagesYamaha Owners Manual 40 50 HPmnbvqwert100% (5)
- BigMike's PH ManifestoDocument31 pagesBigMike's PH ManifestodalbanwaitNo ratings yet
- Enriched Environment Ameliorates DexamethasoneDocument23 pagesEnriched Environment Ameliorates DexamethasoneRoberta RodriguesNo ratings yet
- Article 8 2022 IBRODocument9 pagesArticle 8 2022 IBROEL MOSTAFI HICHAMNo ratings yet
- 1 s2.0 S0278584614001274 MainDocument6 pages1 s2.0 S0278584614001274 MainIkla CavalcanteNo ratings yet
- Main 2Document11 pagesMain 2Jihan MutiaraNo ratings yet
- Social Isolation and Depress Like BehavDocument9 pagesSocial Isolation and Depress Like BehavNityananda PortelladaNo ratings yet
- A Mouse Model of Depression Induced by Repeated Corticosterone InjectionsDocument9 pagesA Mouse Model of Depression Induced by Repeated Corticosterone InjectionsRafiNo ratings yet
- 3 - Maccari, S. - 2014Document17 pages3 - Maccari, S. - 2014João PedroNo ratings yet
- Dalmagro2017 Article MorusNigraAndItsMajorPhenolicSDocument11 pagesDalmagro2017 Article MorusNigraAndItsMajorPhenolicSAna LidiaNo ratings yet
- Synapse Volume 32 Issue 3 1999 (Doi 10.1002/ (Sici) 1098-2396 (19990601) 32:3-187::aid-Syn5-3.0.Co 2-9) Craig W. Berridge Elizabeth Mitton William Clark Robert H. Ro - Engagement in A Non-Escape (DispDocument11 pagesSynapse Volume 32 Issue 3 1999 (Doi 10.1002/ (Sici) 1098-2396 (19990601) 32:3-187::aid-Syn5-3.0.Co 2-9) Craig W. Berridge Elizabeth Mitton William Clark Robert H. Ro - Engagement in A Non-Escape (DispOana IlieNo ratings yet
- Light Dark BoxDocument8 pagesLight Dark Boxattamira95No ratings yet
- Dendritic Reorganization in Pyramidal Neurons in Medial Prefrontal Cortex After Chronic Corticosterone AdministrationDocument9 pagesDendritic Reorganization in Pyramidal Neurons in Medial Prefrontal Cortex After Chronic Corticosterone AdministrationJean Pierre Chastre LuzaNo ratings yet
- Beneficial Effects of Spirulina Platensis, Voluntary Exercise & Environmental Enrichment Against Adolescent Stress InducedDocument12 pagesBeneficial Effects of Spirulina Platensis, Voluntary Exercise & Environmental Enrichment Against Adolescent Stress InducedMARIA PAULA WILCHES ESCOBARNo ratings yet
- 1 s2.0 S0039128X17301204 MainDocument6 pages1 s2.0 S0039128X17301204 MainSab RineNo ratings yet
- Ejercicios Fernando Gomez PinillaDocument14 pagesEjercicios Fernando Gomez PinillaYajaira Saavedra JimenezNo ratings yet
- Zugno2016 PDFDocument34 pagesZugno2016 PDFEka FaridaNo ratings yet
- Glucocorticoid Receptor Regulates Protein Chaperone, Circadian Clock and Affective Disorder Genes in The Zebrafish BrainDocument17 pagesGlucocorticoid Receptor Regulates Protein Chaperone, Circadian Clock and Affective Disorder Genes in The Zebrafish BrainEranElhaikNo ratings yet
- PBB - Repeated Swimming Borsoi Et AlDocument12 pagesPBB - Repeated Swimming Borsoi Et AlPaula LunardiNo ratings yet
- Adipocyte Glucocorticoid Receptors MediateDocument10 pagesAdipocyte Glucocorticoid Receptors MediateRogério Santos SnatusNo ratings yet
- David Et Al 2009Document25 pagesDavid Et Al 2009izaquenNo ratings yet
- Earthing Mat On Stress-Induced Anxiety-Like Behavior and Neuroendocrine Changes in The RatDocument11 pagesEarthing Mat On Stress-Induced Anxiety-Like Behavior and Neuroendocrine Changes in The RatAndre DouradoNo ratings yet
- Stress &healthDocument18 pagesStress &healthsatmayaniNo ratings yet
- 1 s2.0 S0166432805005000 MainDocument9 pages1 s2.0 S0166432805005000 Mainzffkvhtnb7No ratings yet
- Chen Et Al., 2015. Models and Methods To Investigate Acute Stress Responses in CattleDocument28 pagesChen Et Al., 2015. Models and Methods To Investigate Acute Stress Responses in CattleAnahi MedinaNo ratings yet
- 3041-Article Text-13271-1-10-20220414Document7 pages3041-Article Text-13271-1-10-20220414Smonagenes UbNo ratings yet
- Effects of Omega-3 Supplementation On Interleukin and Neurotrophin Levels in An Animal Model of SchizophreniaDocument12 pagesEffects of Omega-3 Supplementation On Interleukin and Neurotrophin Levels in An Animal Model of SchizophreniaIqbal AbdillahNo ratings yet
- Administration of Branched-Chain Amino Acids Alters The Balance Between Pro-Inflammatory and Anti-Inflammatory CytokinesDocument7 pagesAdministration of Branched-Chain Amino Acids Alters The Balance Between Pro-Inflammatory and Anti-Inflammatory CytokinesMatheus PioNo ratings yet
- 1 s2.0 S1056871921001428 MainDocument10 pages1 s2.0 S1056871921001428 Mainsemester6otulNo ratings yet
- Archie, Altmann, Alberts - 2012 - Social Status Predicts Wound Healing in Wild BaboonsDocument11 pagesArchie, Altmann, Alberts - 2012 - Social Status Predicts Wound Healing in Wild BaboonsbenqtenNo ratings yet
- Palmfeldt Et Al. (2016) Protein Biomarkers of Susceptibility and Resilience To Stress in DepressionDocument9 pagesPalmfeldt Et Al. (2016) Protein Biomarkers of Susceptibility and Resilience To Stress in DepressionEdwing Arciniegas CarreñoNo ratings yet
- Antioxidant Treatment AmelioratesDocument26 pagesAntioxidant Treatment Ameliorates2650145987No ratings yet
- Anxiolytic - and Antidepressant-Like Effects of Bacillus Coagulans Unique IS-2 Mediate Via Reshaping of Microbiome Gut-Brain Axis in RatsDocument14 pagesAnxiolytic - and Antidepressant-Like Effects of Bacillus Coagulans Unique IS-2 Mediate Via Reshaping of Microbiome Gut-Brain Axis in RatsJulio QuintanaNo ratings yet
- Aklimatisasi Hewan CobaDocument6 pagesAklimatisasi Hewan CobaNurma FitriaNo ratings yet
- Full PDFDocument9 pagesFull PDFAle TabaskoNo ratings yet
- A Diet High in Fat and Sugar Reverses Anxiety-Like Behaviour Induced by Limited Nesting in Male RatsDocument8 pagesA Diet High in Fat and Sugar Reverses Anxiety-Like Behaviour Induced by Limited Nesting in Male RatsAldehydeNo ratings yet
- FullDocument13 pagesFullAulas EspañolNo ratings yet
- Er Stress DissertationDocument5 pagesEr Stress DissertationCustomCollegePapersCanada100% (1)
- 1 s2.0 S2352289523000899 MainDocument10 pages1 s2.0 S2352289523000899 Maincamilobermudez.pinturasmundialNo ratings yet
- DHEA DR TamilDocument4 pagesDHEA DR TamilDr. Santhi Silambanan, Dept. of Biochemistry, SRUNo ratings yet
- 2015.05 La Ingesta de Prebióticos Reduce La Respuesta de Cortisol Al Despertar y Altera El Sesgo Emocional en Voluntarios SanosDocument9 pages2015.05 La Ingesta de Prebióticos Reduce La Respuesta de Cortisol Al Despertar y Altera El Sesgo Emocional en Voluntarios SanosEdward UriarteNo ratings yet
- Stress Control and Human Nutrition: ReviewDocument7 pagesStress Control and Human Nutrition: Reviewgiulia.santinNo ratings yet
- Early-Life Adversity and Long-Term VaisermanDocument16 pagesEarly-Life Adversity and Long-Term VaisermanJennyNo ratings yet
- Hup 2379Document12 pagesHup 2379Isabely AmabilyNo ratings yet
- Fnbeh 15 700182Document15 pagesFnbeh 15 700182samawi ramudNo ratings yet
- Apha 13440Document17 pagesApha 13440javillusNo ratings yet
- Maiaoliveira2021 - Neuroprotective and Antioxidant Effects ofDocument11 pagesMaiaoliveira2021 - Neuroprotective and Antioxidant Effects ofRAQUEL CAMELO BARRETO DE SIQUEIRANo ratings yet
- Transferring The Blues Depression Associated Gut Microbiota Induces Neurobehavioural Changes in The Rat.Document11 pagesTransferring The Blues Depression Associated Gut Microbiota Induces Neurobehavioural Changes in The Rat.Dori BearNo ratings yet
- Psychobiology and Molecular Genetics of ResilienceDocument23 pagesPsychobiology and Molecular Genetics of Resiliencemelpomena81No ratings yet
- Stress and The Gastrointestinal Tract: ReviewDocument8 pagesStress and The Gastrointestinal Tract: ReviewsufaNo ratings yet
- 17-Estradiol Attenuates Hippocampal Neuronal LossDocument9 pages17-Estradiol Attenuates Hippocampal Neuronal LossrodrigounitedNo ratings yet
- Acute and long-term effects of intracerebroventricular administration of α-ketoisocaproic acid on oxidative stress parameters and cognitive and noncognitive behaviorsDocument12 pagesAcute and long-term effects of intracerebroventricular administration of α-ketoisocaproic acid on oxidative stress parameters and cognitive and noncognitive behaviorsMatheus PioNo ratings yet
- Hyperglycemic Effect On Brain Cholinergic Functions, Oxidative Stress and Protein Expression of Brain Derived Neurotropic Factor (BDNF) On Cognitive Functions in Streptozotocin Induced-Diabetic RatsDocument9 pagesHyperglycemic Effect On Brain Cholinergic Functions, Oxidative Stress and Protein Expression of Brain Derived Neurotropic Factor (BDNF) On Cognitive Functions in Streptozotocin Induced-Diabetic Ratsd21m19No ratings yet
- Chinese Phytotherapy To Reduce Stress, Anxiety and Improve Quality of Life: Randomized Controlled TrialDocument8 pagesChinese Phytotherapy To Reduce Stress, Anxiety and Improve Quality of Life: Randomized Controlled Trialthunder.egeNo ratings yet
- Estrés Crónico, Obesidad Visceral. Kyrou y TsigosDocument7 pagesEstrés Crónico, Obesidad Visceral. Kyrou y TsigosBenja MorenoNo ratings yet
- PETA Letter To HarvardDocument4 pagesPETA Letter To HarvardWashington ExaminerNo ratings yet
- Sexual Dimorphism Spatial Learning Metabolism Stress Diet (10-14)Document5 pagesSexual Dimorphism Spatial Learning Metabolism Stress Diet (10-14)Camila Alejandra Inostroza RiveraNo ratings yet
- Stress Hormones Physiological Stress andDocument7 pagesStress Hormones Physiological Stress andA1 [2] ضرغام لطيف سلمان داود / مسائيNo ratings yet
- 2016 Article 771Document14 pages2016 Article 771Büşra YılmazNo ratings yet
- Artigo Desmame PrecoceDocument10 pagesArtigo Desmame Precocekayllane.vasconcelosNo ratings yet
- Journal of Neuroimmunology: SciencedirectDocument10 pagesJournal of Neuroimmunology: SciencedirectPsiquiatria TranslacionalNo ratings yet
- Gale Researcher Guide for: Physical and Chemical Bases of Stress and CopingFrom EverandGale Researcher Guide for: Physical and Chemical Bases of Stress and CopingNo ratings yet
- The Economics of Community Forest Management in Madagascar: Is There A Free Lunch? - 2007Document97 pagesThe Economics of Community Forest Management in Madagascar: Is There A Free Lunch? - 2007HayZara MadagascarNo ratings yet
- MAR 20 2Q15:: Department of Public Works and HighwaysDocument6 pagesMAR 20 2Q15:: Department of Public Works and HighwaysFaustino AbadNo ratings yet
- Military Gear Suppliers Expo Attendees.Document32 pagesMilitary Gear Suppliers Expo Attendees.Waf EtanoNo ratings yet
- Test - R.A 9514 Fire Code - Quizlet PDFDocument3 pagesTest - R.A 9514 Fire Code - Quizlet PDFPenuel G. Bantog100% (1)
- Narrative Letter Additional AdditionalDocument3 pagesNarrative Letter Additional AdditionalIUSECAPSLOCKNo ratings yet
- Heartfulness Magazine - January 2022 (Volume 7, Issue 1)Document84 pagesHeartfulness Magazine - January 2022 (Volume 7, Issue 1)HeartfulnessNo ratings yet
- UnconsciousnessDocument6 pagesUnconsciousnessSeema RahulNo ratings yet
- Development of A Smart Garment For The Assessment of Cardiac Mechanical Performance and Other Vital Signs During Sleep in MicrogravityDocument9 pagesDevelopment of A Smart Garment For The Assessment of Cardiac Mechanical Performance and Other Vital Signs During Sleep in MicrogravitynarykNo ratings yet
- Hartley 1980Document8 pagesHartley 1980Alexes Paul TancioNo ratings yet
- Practical Manual Hort 382Document112 pagesPractical Manual Hort 382Dr.Eswara Reddy Siddareddy100% (1)
- Post Assesment Question 1Document7 pagesPost Assesment Question 1Ganesh GuptaNo ratings yet
- Boronizing AVIONDocument37 pagesBoronizing AVIONManwi Khandelwal100% (1)
- The Economic and Security Intricacies of The BangsamoroDocument19 pagesThe Economic and Security Intricacies of The BangsamoroSigmund Alfred Ducusin100% (2)
- STart Max Brochure EN - CompressedDocument4 pagesSTart Max Brochure EN - Compressedb4753485No ratings yet
- CS1354 - Graphics and Mutimedia Important Two MarksDocument17 pagesCS1354 - Graphics and Mutimedia Important Two MarksSumathi BasNo ratings yet
- Atm-1022 Mechanical Workshop Module 3 PDFDocument19 pagesAtm-1022 Mechanical Workshop Module 3 PDFMohamed Sayed Abdel GaffarNo ratings yet
- Generador Perkins A65pe3Document4 pagesGenerador Perkins A65pe3cauditarosalesNo ratings yet
- Bernal Et Al. - 2011 - Advanced Analysis of NutraceuticalsDocument17 pagesBernal Et Al. - 2011 - Advanced Analysis of NutraceuticalsAaron Quispe ChambiNo ratings yet
- Learnenglish Podcasts Elementary 02 10 Support PackDocument19 pagesLearnenglish Podcasts Elementary 02 10 Support PackWilliams NiñoNo ratings yet
- Muhammad Dzaki Fadhilah-195020307111053-EXERCISE 14 Multiple Linear RegressionDocument24 pagesMuhammad Dzaki Fadhilah-195020307111053-EXERCISE 14 Multiple Linear Regressionazka rikiNo ratings yet
- We Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsDocument19 pagesWe Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsBendaud bataborNo ratings yet
- M Tech SP ADSD Model Question PaperDocument2 pagesM Tech SP ADSD Model Question Papersafu_117No ratings yet
- Power Plant Engineering NotesDocument3 pagesPower Plant Engineering NotesNeelesh PandeyNo ratings yet
- Iteration Question'sDocument17 pagesIteration Question'sAyra MujibNo ratings yet
- Development of A Two-Wheeled Mobile Tilting & Balancing (MTB) RobotDocument6 pagesDevelopment of A Two-Wheeled Mobile Tilting & Balancing (MTB) RobotAs'ad Syamsul ArifinNo ratings yet
- Square Portal FrameDocument8 pagesSquare Portal FramenabilaNo ratings yet
- Data Sheet ST ESTABILIZADORDocument3 pagesData Sheet ST ESTABILIZADORJuan Antonio Barco MorenoNo ratings yet



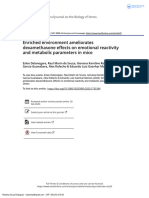





















































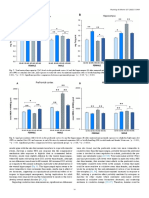
































Download as pdf or txt
You might also like
- Task Risk Assessment For RadiographyDocument5 pagesTask Risk Assessment For RadiographySanjeev Nair81% (21)
- Yamaha Owners Manual 40 50 HPDocument88 pagesYamaha Owners Manual 40 50 HPmnbvqwert100% (5)
- BigMike's PH ManifestoDocument31 pagesBigMike's PH ManifestodalbanwaitNo ratings yet
- Enriched Environment Ameliorates DexamethasoneDocument23 pagesEnriched Environment Ameliorates DexamethasoneRoberta RodriguesNo ratings yet
- Article 8 2022 IBRODocument9 pagesArticle 8 2022 IBROEL MOSTAFI HICHAMNo ratings yet
- 1 s2.0 S0278584614001274 MainDocument6 pages1 s2.0 S0278584614001274 MainIkla CavalcanteNo ratings yet
- Main 2Document11 pagesMain 2Jihan MutiaraNo ratings yet
- Social Isolation and Depress Like BehavDocument9 pagesSocial Isolation and Depress Like BehavNityananda PortelladaNo ratings yet
- A Mouse Model of Depression Induced by Repeated Corticosterone InjectionsDocument9 pagesA Mouse Model of Depression Induced by Repeated Corticosterone InjectionsRafiNo ratings yet
- 3 - Maccari, S. - 2014Document17 pages3 - Maccari, S. - 2014João PedroNo ratings yet
- Dalmagro2017 Article MorusNigraAndItsMajorPhenolicSDocument11 pagesDalmagro2017 Article MorusNigraAndItsMajorPhenolicSAna LidiaNo ratings yet
- Synapse Volume 32 Issue 3 1999 (Doi 10.1002/ (Sici) 1098-2396 (19990601) 32:3-187::aid-Syn5-3.0.Co 2-9) Craig W. Berridge Elizabeth Mitton William Clark Robert H. Ro - Engagement in A Non-Escape (DispDocument11 pagesSynapse Volume 32 Issue 3 1999 (Doi 10.1002/ (Sici) 1098-2396 (19990601) 32:3-187::aid-Syn5-3.0.Co 2-9) Craig W. Berridge Elizabeth Mitton William Clark Robert H. Ro - Engagement in A Non-Escape (DispOana IlieNo ratings yet
- Light Dark BoxDocument8 pagesLight Dark Boxattamira95No ratings yet
- Dendritic Reorganization in Pyramidal Neurons in Medial Prefrontal Cortex After Chronic Corticosterone AdministrationDocument9 pagesDendritic Reorganization in Pyramidal Neurons in Medial Prefrontal Cortex After Chronic Corticosterone AdministrationJean Pierre Chastre LuzaNo ratings yet
- Beneficial Effects of Spirulina Platensis, Voluntary Exercise & Environmental Enrichment Against Adolescent Stress InducedDocument12 pagesBeneficial Effects of Spirulina Platensis, Voluntary Exercise & Environmental Enrichment Against Adolescent Stress InducedMARIA PAULA WILCHES ESCOBARNo ratings yet
- 1 s2.0 S0039128X17301204 MainDocument6 pages1 s2.0 S0039128X17301204 MainSab RineNo ratings yet
- Ejercicios Fernando Gomez PinillaDocument14 pagesEjercicios Fernando Gomez PinillaYajaira Saavedra JimenezNo ratings yet
- Zugno2016 PDFDocument34 pagesZugno2016 PDFEka FaridaNo ratings yet
- Glucocorticoid Receptor Regulates Protein Chaperone, Circadian Clock and Affective Disorder Genes in The Zebrafish BrainDocument17 pagesGlucocorticoid Receptor Regulates Protein Chaperone, Circadian Clock and Affective Disorder Genes in The Zebrafish BrainEranElhaikNo ratings yet
- PBB - Repeated Swimming Borsoi Et AlDocument12 pagesPBB - Repeated Swimming Borsoi Et AlPaula LunardiNo ratings yet
- Adipocyte Glucocorticoid Receptors MediateDocument10 pagesAdipocyte Glucocorticoid Receptors MediateRogério Santos SnatusNo ratings yet
- David Et Al 2009Document25 pagesDavid Et Al 2009izaquenNo ratings yet
- Earthing Mat On Stress-Induced Anxiety-Like Behavior and Neuroendocrine Changes in The RatDocument11 pagesEarthing Mat On Stress-Induced Anxiety-Like Behavior and Neuroendocrine Changes in The RatAndre DouradoNo ratings yet
- Stress &healthDocument18 pagesStress &healthsatmayaniNo ratings yet
- 1 s2.0 S0166432805005000 MainDocument9 pages1 s2.0 S0166432805005000 Mainzffkvhtnb7No ratings yet
- Chen Et Al., 2015. Models and Methods To Investigate Acute Stress Responses in CattleDocument28 pagesChen Et Al., 2015. Models and Methods To Investigate Acute Stress Responses in CattleAnahi MedinaNo ratings yet
- 3041-Article Text-13271-1-10-20220414Document7 pages3041-Article Text-13271-1-10-20220414Smonagenes UbNo ratings yet
- Effects of Omega-3 Supplementation On Interleukin and Neurotrophin Levels in An Animal Model of SchizophreniaDocument12 pagesEffects of Omega-3 Supplementation On Interleukin and Neurotrophin Levels in An Animal Model of SchizophreniaIqbal AbdillahNo ratings yet
- Administration of Branched-Chain Amino Acids Alters The Balance Between Pro-Inflammatory and Anti-Inflammatory CytokinesDocument7 pagesAdministration of Branched-Chain Amino Acids Alters The Balance Between Pro-Inflammatory and Anti-Inflammatory CytokinesMatheus PioNo ratings yet
- 1 s2.0 S1056871921001428 MainDocument10 pages1 s2.0 S1056871921001428 Mainsemester6otulNo ratings yet
- Archie, Altmann, Alberts - 2012 - Social Status Predicts Wound Healing in Wild BaboonsDocument11 pagesArchie, Altmann, Alberts - 2012 - Social Status Predicts Wound Healing in Wild BaboonsbenqtenNo ratings yet
- Palmfeldt Et Al. (2016) Protein Biomarkers of Susceptibility and Resilience To Stress in DepressionDocument9 pagesPalmfeldt Et Al. (2016) Protein Biomarkers of Susceptibility and Resilience To Stress in DepressionEdwing Arciniegas CarreñoNo ratings yet
- Antioxidant Treatment AmelioratesDocument26 pagesAntioxidant Treatment Ameliorates2650145987No ratings yet
- Anxiolytic - and Antidepressant-Like Effects of Bacillus Coagulans Unique IS-2 Mediate Via Reshaping of Microbiome Gut-Brain Axis in RatsDocument14 pagesAnxiolytic - and Antidepressant-Like Effects of Bacillus Coagulans Unique IS-2 Mediate Via Reshaping of Microbiome Gut-Brain Axis in RatsJulio QuintanaNo ratings yet
- Aklimatisasi Hewan CobaDocument6 pagesAklimatisasi Hewan CobaNurma FitriaNo ratings yet
- Full PDFDocument9 pagesFull PDFAle TabaskoNo ratings yet
- A Diet High in Fat and Sugar Reverses Anxiety-Like Behaviour Induced by Limited Nesting in Male RatsDocument8 pagesA Diet High in Fat and Sugar Reverses Anxiety-Like Behaviour Induced by Limited Nesting in Male RatsAldehydeNo ratings yet
- FullDocument13 pagesFullAulas EspañolNo ratings yet
- Er Stress DissertationDocument5 pagesEr Stress DissertationCustomCollegePapersCanada100% (1)
- 1 s2.0 S2352289523000899 MainDocument10 pages1 s2.0 S2352289523000899 Maincamilobermudez.pinturasmundialNo ratings yet
- DHEA DR TamilDocument4 pagesDHEA DR TamilDr. Santhi Silambanan, Dept. of Biochemistry, SRUNo ratings yet
- 2015.05 La Ingesta de Prebióticos Reduce La Respuesta de Cortisol Al Despertar y Altera El Sesgo Emocional en Voluntarios SanosDocument9 pages2015.05 La Ingesta de Prebióticos Reduce La Respuesta de Cortisol Al Despertar y Altera El Sesgo Emocional en Voluntarios SanosEdward UriarteNo ratings yet
- Stress Control and Human Nutrition: ReviewDocument7 pagesStress Control and Human Nutrition: Reviewgiulia.santinNo ratings yet
- Early-Life Adversity and Long-Term VaisermanDocument16 pagesEarly-Life Adversity and Long-Term VaisermanJennyNo ratings yet
- Hup 2379Document12 pagesHup 2379Isabely AmabilyNo ratings yet
- Fnbeh 15 700182Document15 pagesFnbeh 15 700182samawi ramudNo ratings yet
- Apha 13440Document17 pagesApha 13440javillusNo ratings yet
- Maiaoliveira2021 - Neuroprotective and Antioxidant Effects ofDocument11 pagesMaiaoliveira2021 - Neuroprotective and Antioxidant Effects ofRAQUEL CAMELO BARRETO DE SIQUEIRANo ratings yet
- Transferring The Blues Depression Associated Gut Microbiota Induces Neurobehavioural Changes in The Rat.Document11 pagesTransferring The Blues Depression Associated Gut Microbiota Induces Neurobehavioural Changes in The Rat.Dori BearNo ratings yet
- Psychobiology and Molecular Genetics of ResilienceDocument23 pagesPsychobiology and Molecular Genetics of Resiliencemelpomena81No ratings yet
- Stress and The Gastrointestinal Tract: ReviewDocument8 pagesStress and The Gastrointestinal Tract: ReviewsufaNo ratings yet
- 17-Estradiol Attenuates Hippocampal Neuronal LossDocument9 pages17-Estradiol Attenuates Hippocampal Neuronal LossrodrigounitedNo ratings yet
- Acute and long-term effects of intracerebroventricular administration of α-ketoisocaproic acid on oxidative stress parameters and cognitive and noncognitive behaviorsDocument12 pagesAcute and long-term effects of intracerebroventricular administration of α-ketoisocaproic acid on oxidative stress parameters and cognitive and noncognitive behaviorsMatheus PioNo ratings yet
- Hyperglycemic Effect On Brain Cholinergic Functions, Oxidative Stress and Protein Expression of Brain Derived Neurotropic Factor (BDNF) On Cognitive Functions in Streptozotocin Induced-Diabetic RatsDocument9 pagesHyperglycemic Effect On Brain Cholinergic Functions, Oxidative Stress and Protein Expression of Brain Derived Neurotropic Factor (BDNF) On Cognitive Functions in Streptozotocin Induced-Diabetic Ratsd21m19No ratings yet
- Chinese Phytotherapy To Reduce Stress, Anxiety and Improve Quality of Life: Randomized Controlled TrialDocument8 pagesChinese Phytotherapy To Reduce Stress, Anxiety and Improve Quality of Life: Randomized Controlled Trialthunder.egeNo ratings yet
- Estrés Crónico, Obesidad Visceral. Kyrou y TsigosDocument7 pagesEstrés Crónico, Obesidad Visceral. Kyrou y TsigosBenja MorenoNo ratings yet
- PETA Letter To HarvardDocument4 pagesPETA Letter To HarvardWashington ExaminerNo ratings yet
- Sexual Dimorphism Spatial Learning Metabolism Stress Diet (10-14)Document5 pagesSexual Dimorphism Spatial Learning Metabolism Stress Diet (10-14)Camila Alejandra Inostroza RiveraNo ratings yet
- Stress Hormones Physiological Stress andDocument7 pagesStress Hormones Physiological Stress andA1 [2] ضرغام لطيف سلمان داود / مسائيNo ratings yet
- 2016 Article 771Document14 pages2016 Article 771Büşra YılmazNo ratings yet
- Artigo Desmame PrecoceDocument10 pagesArtigo Desmame Precocekayllane.vasconcelosNo ratings yet
- Journal of Neuroimmunology: SciencedirectDocument10 pagesJournal of Neuroimmunology: SciencedirectPsiquiatria TranslacionalNo ratings yet
- Gale Researcher Guide for: Physical and Chemical Bases of Stress and CopingFrom EverandGale Researcher Guide for: Physical and Chemical Bases of Stress and CopingNo ratings yet
- The Economics of Community Forest Management in Madagascar: Is There A Free Lunch? - 2007Document97 pagesThe Economics of Community Forest Management in Madagascar: Is There A Free Lunch? - 2007HayZara MadagascarNo ratings yet
- MAR 20 2Q15:: Department of Public Works and HighwaysDocument6 pagesMAR 20 2Q15:: Department of Public Works and HighwaysFaustino AbadNo ratings yet
- Military Gear Suppliers Expo Attendees.Document32 pagesMilitary Gear Suppliers Expo Attendees.Waf EtanoNo ratings yet
- Test - R.A 9514 Fire Code - Quizlet PDFDocument3 pagesTest - R.A 9514 Fire Code - Quizlet PDFPenuel G. Bantog100% (1)
- Narrative Letter Additional AdditionalDocument3 pagesNarrative Letter Additional AdditionalIUSECAPSLOCKNo ratings yet
- Heartfulness Magazine - January 2022 (Volume 7, Issue 1)Document84 pagesHeartfulness Magazine - January 2022 (Volume 7, Issue 1)HeartfulnessNo ratings yet
- UnconsciousnessDocument6 pagesUnconsciousnessSeema RahulNo ratings yet
- Development of A Smart Garment For The Assessment of Cardiac Mechanical Performance and Other Vital Signs During Sleep in MicrogravityDocument9 pagesDevelopment of A Smart Garment For The Assessment of Cardiac Mechanical Performance and Other Vital Signs During Sleep in MicrogravitynarykNo ratings yet
- Hartley 1980Document8 pagesHartley 1980Alexes Paul TancioNo ratings yet
- Practical Manual Hort 382Document112 pagesPractical Manual Hort 382Dr.Eswara Reddy Siddareddy100% (1)
- Post Assesment Question 1Document7 pagesPost Assesment Question 1Ganesh GuptaNo ratings yet
- Boronizing AVIONDocument37 pagesBoronizing AVIONManwi Khandelwal100% (1)
- The Economic and Security Intricacies of The BangsamoroDocument19 pagesThe Economic and Security Intricacies of The BangsamoroSigmund Alfred Ducusin100% (2)
- STart Max Brochure EN - CompressedDocument4 pagesSTart Max Brochure EN - Compressedb4753485No ratings yet
- CS1354 - Graphics and Mutimedia Important Two MarksDocument17 pagesCS1354 - Graphics and Mutimedia Important Two MarksSumathi BasNo ratings yet
- Atm-1022 Mechanical Workshop Module 3 PDFDocument19 pagesAtm-1022 Mechanical Workshop Module 3 PDFMohamed Sayed Abdel GaffarNo ratings yet
- Generador Perkins A65pe3Document4 pagesGenerador Perkins A65pe3cauditarosalesNo ratings yet
- Bernal Et Al. - 2011 - Advanced Analysis of NutraceuticalsDocument17 pagesBernal Et Al. - 2011 - Advanced Analysis of NutraceuticalsAaron Quispe ChambiNo ratings yet
- Learnenglish Podcasts Elementary 02 10 Support PackDocument19 pagesLearnenglish Podcasts Elementary 02 10 Support PackWilliams NiñoNo ratings yet
- Muhammad Dzaki Fadhilah-195020307111053-EXERCISE 14 Multiple Linear RegressionDocument24 pagesMuhammad Dzaki Fadhilah-195020307111053-EXERCISE 14 Multiple Linear Regressionazka rikiNo ratings yet
- We Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsDocument19 pagesWe Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsBendaud bataborNo ratings yet
- M Tech SP ADSD Model Question PaperDocument2 pagesM Tech SP ADSD Model Question Papersafu_117No ratings yet
- Power Plant Engineering NotesDocument3 pagesPower Plant Engineering NotesNeelesh PandeyNo ratings yet
- Iteration Question'sDocument17 pagesIteration Question'sAyra MujibNo ratings yet
- Development of A Two-Wheeled Mobile Tilting & Balancing (MTB) RobotDocument6 pagesDevelopment of A Two-Wheeled Mobile Tilting & Balancing (MTB) RobotAs'ad Syamsul ArifinNo ratings yet
- Square Portal FrameDocument8 pagesSquare Portal FramenabilaNo ratings yet
- Data Sheet ST ESTABILIZADORDocument3 pagesData Sheet ST ESTABILIZADORJuan Antonio Barco MorenoNo ratings yet