Download as pdf or txt
You might also like
- The Stabilizing System of The Spine Part 1..panjabiDocument7 pagesThe Stabilizing System of The Spine Part 1..panjabilgminassianNo ratings yet
- Abstract BookDocument213 pagesAbstract BookSherry HeinzNo ratings yet
- Transcranial Magnetic StimulationDocument2 pagesTranscranial Magnetic StimulationCandle Shop FullNo ratings yet
- New Targets For RTMS in Depression - Review of Convergent Evidence - 2013Document10 pagesNew Targets For RTMS in Depression - Review of Convergent Evidence - 2013Lina SantosNo ratings yet
- Peterka - 2002 - J NeurophysDocument23 pagesPeterka - 2002 - J NeurophysKarem TuctoNo ratings yet
- An Exploration of The Function of The Triceps Surae During Normal Gait Using Functional Electrical Stimulation PDFDocument2 pagesAn Exploration of The Function of The Triceps Surae During Normal Gait Using Functional Electrical Stimulation PDFValentinus Adi PamungkasNo ratings yet
- Eight Weeks of Proprioceptive Neuromuscular FaciliDocument9 pagesEight Weeks of Proprioceptive Neuromuscular FaciliMauricio MoncadaNo ratings yet
- 1 s2.0 S1388245704000975 MainDocument12 pages1 s2.0 S1388245704000975 MainpashaNo ratings yet
- Applied Kinesiology Essentials DR S Cuthbert Townsend Letter Jan.2016 PDFDocument5 pagesApplied Kinesiology Essentials DR S Cuthbert Townsend Letter Jan.2016 PDFdocerick87No ratings yet
- Fnint 11 00002Document15 pagesFnint 11 00002labsoneducationNo ratings yet
- Yulandy ArticuloDocument7 pagesYulandy ArticuloLaura Carolina Pabon SandovalNo ratings yet
- Ozer 2006 Neuromuscular Electrical StimulatioDocument5 pagesOzer 2006 Neuromuscular Electrical StimulatioAmr Mohamed GalalNo ratings yet
- Muscle Fatigue: What, Why and How It Influences Muscle FunctionDocument13 pagesMuscle Fatigue: What, Why and How It Influences Muscle FunctionAna Marta AnastácioNo ratings yet
- Ates F Et Al 2019Document10 pagesAtes F Et Al 2019blaiNo ratings yet
- Abboud Et Al. - 2016 - Influence of Lumbar Muscle Fatigue On Trunk AdaptaDocument16 pagesAbboud Et Al. - 2016 - Influence of Lumbar Muscle Fatigue On Trunk AdaptaYouriDuchêneNo ratings yet
- Paper Positional Release Technique SF MWDocument17 pagesPaper Positional Release Technique SF MWkamNo ratings yet
- Pearce 2012Document9 pagesPearce 2012edu.10038No ratings yet
- Effects of Abdominal Stabilization ManeuDocument12 pagesEffects of Abdominal Stabilization ManeuMarcos BurónNo ratings yet
- Spasticity: Pathophysiology and Neural Control: Simon M. Danner, Milan R. DimitrijevicDocument6 pagesSpasticity: Pathophysiology and Neural Control: Simon M. Danner, Milan R. DimitrijevicDimas Gatra DiantoroNo ratings yet
- Vera Garcia 2006 Effects of Different Levels of Torso Coactivation On Trunk Muscular and Kinematic Responses To PoDocument13 pagesVera Garcia 2006 Effects of Different Levels of Torso Coactivation On Trunk Muscular and Kinematic Responses To PoMichael JunNo ratings yet
- Rossi2014 PDFDocument8 pagesRossi2014 PDFDiana Magnavita MouraNo ratings yet
- Chronic Resistance Training Is It Time To RethinkDocument11 pagesChronic Resistance Training Is It Time To RethinkDr. Andre Lopes PhD.No ratings yet
- Evaluating The Effects of Stress Reduction Exercises Employing Mild Tremors: A Pilot StudyDocument23 pagesEvaluating The Effects of Stress Reduction Exercises Employing Mild Tremors: A Pilot StudydNo ratings yet
- ArtigoDocument10 pagesArtigoLevyNo ratings yet
- Adaptive Responses To Muscle Lengthening and Shortening in HumansDocument8 pagesAdaptive Responses To Muscle Lengthening and Shortening in Humansrafaeldfonseca09No ratings yet
- Biomechanism of StretchingDocument11 pagesBiomechanism of Stretchingsara mohamedNo ratings yet
- Mohamed Abdalla 211080013Document11 pagesMohamed Abdalla 211080013rehab aymanNo ratings yet
- Muscle Fatigue What Why and How It Influences MuscDocument14 pagesMuscle Fatigue What Why and How It Influences MuscWeCareNo ratings yet
- Effects of Abdominal Stabilization ManeuDocument12 pagesEffects of Abdominal Stabilization ManeuMarcos BurónNo ratings yet
- 2002 Peterka - Sensoriomotor Integration in Human Postural ControlDocument23 pages2002 Peterka - Sensoriomotor Integration in Human Postural ControlLoreto GálvezNo ratings yet
- 2013 Shape Shifting PainDocument18 pages2013 Shape Shifting PainCristian OyarzoNo ratings yet
- Muscle Tone and JoinDocument12 pagesMuscle Tone and JoinomarihuanoNo ratings yet
- Kidgell Et Al-2017-European Journal of NeuroscienceDocument14 pagesKidgell Et Al-2017-European Journal of NeurosciencePedro Alves FernandesNo ratings yet
- Muscle FatigueDocument13 pagesMuscle FatigueNadiyah Abudan100% (1)
- Human Postural ControlDocument9 pagesHuman Postural Controlelbueno21No ratings yet
- Objective Evaluation of A Splint Designed To Reduce H YpertonicityDocument6 pagesObjective Evaluation of A Splint Designed To Reduce H YpertonicityNoor efNo ratings yet
- LatellaPearceKidgel EJAPDocument11 pagesLatellaPearceKidgel EJAPMarcos BurónNo ratings yet
- Muscle Tone PhysiologyDocument21 pagesMuscle Tone PhysiologySagar SangoleNo ratings yet
- The Biomechanics of Stretching: Duane KnudsonDocument10 pagesThe Biomechanics of Stretching: Duane KnudsonAnuragNo ratings yet
- The Biomechanics of Stretching: Duane KnudsonDocument10 pagesThe Biomechanics of Stretching: Duane KnudsonSanjanaNo ratings yet
- Mental Pratice With Motor Imagery in Strock RecoveryDocument14 pagesMental Pratice With Motor Imagery in Strock RecoverygeohawaiiNo ratings yet
- Stretch-Induced Enhancement of Mechanical PowerDocument8 pagesStretch-Induced Enhancement of Mechanical PowerCharisNo ratings yet
- Barss 2020Document14 pagesBarss 2020edu.10038No ratings yet
- KSPM 12 02 121Document8 pagesKSPM 12 02 121kang soon cheolNo ratings yet
- Manual Therapy: Hiroshi Takasaki, Takeshi Iizawa, Toby Hall, Takuo Nakamura, Shouta KanekoDocument6 pagesManual Therapy: Hiroshi Takasaki, Takeshi Iizawa, Toby Hall, Takuo Nakamura, Shouta KanekoDiego Cruces OrdoñezNo ratings yet
- Brain: Altering Spinal Cord Excitability Enables Voluntary Movements After Chronic Complete Paralysis in HumansDocument16 pagesBrain: Altering Spinal Cord Excitability Enables Voluntary Movements After Chronic Complete Paralysis in HumansSebastián GallegosNo ratings yet
- Journal of Bodywork & Movement TherapiesDocument6 pagesJournal of Bodywork & Movement TherapiesLuqmanul HakimNo ratings yet
- Piis 1360859215001679Document6 pagesPiis 1360859215001679api-318417723No ratings yet
- Clinical Neurodynamics and Sports Medicine: Origins and DevelopmentDocument8 pagesClinical Neurodynamics and Sports Medicine: Origins and DevelopmentWahid NasrudinNo ratings yet
- Investigating Interindividual Variability in Corticomotor Reorganization During Sustained Hamstring Pain: A Randomized Experimental StudyDocument12 pagesInvestigating Interindividual Variability in Corticomotor Reorganization During Sustained Hamstring Pain: A Randomized Experimental StudyJosé Pedro CorreiaNo ratings yet
- Yong-Hyun 2011Document7 pagesYong-Hyun 2011GorkaBuesaNo ratings yet
- 28789-Ce (Ra1) F (GG) PF1 (Aa SS) Pfa (MJ GG) PF2 (MJ SS)Document4 pages28789-Ce (Ra1) F (GG) PF1 (Aa SS) Pfa (MJ GG) PF2 (MJ SS)vinika ChaudharyNo ratings yet
- Neck Proprioception Shapes Body Orientation and Perception of MotionDocument13 pagesNeck Proprioception Shapes Body Orientation and Perception of Motion伊文超100% (1)
- Posture Control AND Trajectory Formation During ARM Movement'Document7 pagesPosture Control AND Trajectory Formation During ARM Movement'Krishna VeniNo ratings yet
- Hynes and Turner (2020)Document10 pagesHynes and Turner (2020)Marzio FagioliNo ratings yet
- Estabilizacion Espinal P1Document5 pagesEstabilizacion Espinal P1Jenny IldefonsoNo ratings yet
- 2009 Ericsson Et Al-2009-Scandinavian Journal of Medicine & Science in SportsDocument10 pages2009 Ericsson Et Al-2009-Scandinavian Journal of Medicine & Science in SportsLuis MiguelNo ratings yet
- Frog 2005Document11 pagesFrog 2005Alfi OktafaniNo ratings yet
- Trappe 2004Document13 pagesTrappe 2004cristianNo ratings yet
- Muscle Atrophy in Patients Receiving Hemodialysis: Effects On Muscle Strength, Muscle Quality, and Physical FunctionDocument7 pagesMuscle Atrophy in Patients Receiving Hemodialysis: Effects On Muscle Strength, Muscle Quality, and Physical FunctionPaulo GomesNo ratings yet
- Mitchell Et Al. (2009) Reflex Mechanisms Lack of ContributionDocument16 pagesMitchell Et Al. (2009) Reflex Mechanisms Lack of ContributionRafael RodriguesNo ratings yet
- Sensorimotor Training A Global Approach For BalancDocument9 pagesSensorimotor Training A Global Approach For BalancVizaNo ratings yet
- Teach Yourself Pali in EnglishDocument252 pagesTeach Yourself Pali in EnglishHuyentrang VuNo ratings yet
- My Understanding of Asubha Practice - Ajahn Brahmavamso, 2001, (16p)Document16 pagesMy Understanding of Asubha Practice - Ajahn Brahmavamso, 2001, (16p)Huyentrang VuNo ratings yet
- Functional Neuroanatomy of Meditation (78 Functional Neuroimaging Investigations) - Kieran C.R. Fox, 2016 (21p)Document21 pagesFunctional Neuroanatomy of Meditation (78 Functional Neuroimaging Investigations) - Kieran C.R. Fox, 2016 (21p)Huyentrang VuNo ratings yet
- How To Live As A Good Buddhist, Vol.1 - U Han Htay, Yangon-2002 (575p)Document575 pagesHow To Live As A Good Buddhist, Vol.1 - U Han Htay, Yangon-2002 (575p)Huyentrang VuNo ratings yet
- VMC 05 enDocument16 pagesVMC 05 enHuyentrang VuNo ratings yet
- VMC 02 enDocument12 pagesVMC 02 enHuyentrang VuNo ratings yet
- The Way of The Ultimate Goal (Nibbana) in Mangala Sutta, Vol.5, Issue 3 - Chandadhika, Mylapore-2015 (10p)Document10 pagesThe Way of The Ultimate Goal (Nibbana) in Mangala Sutta, Vol.5, Issue 3 - Chandadhika, Mylapore-2015 (10p)Huyentrang VuNo ratings yet
- The Immediate and Long-Term Effects of An Intensive Meditation Retreat - J.N. Cohen Et Al., (PRT 1064-77 Mindfulness 8) CrossMark-2017 (14p)Document14 pagesThe Immediate and Long-Term Effects of An Intensive Meditation Retreat - J.N. Cohen Et Al., (PRT 1064-77 Mindfulness 8) CrossMark-2017 (14p)Huyentrang VuNo ratings yet
- Personality and Individual Di FferencesDocument4 pagesPersonality and Individual Di FferencesHuyentrang VuNo ratings yet
- Comparing and Contrasting The Theravadin Conception of Anatta With Sunyata (Mahayana Concepts, Daoist Ideas) - Shaun Ramsden (26p)Document26 pagesComparing and Contrasting The Theravadin Conception of Anatta With Sunyata (Mahayana Concepts, Daoist Ideas) - Shaun Ramsden (26p)Huyentrang VuNo ratings yet
- IWP21 01 DiufDocument9 pagesIWP21 01 DiufSai Vamshi PranayNo ratings yet
- Materi Iht TMS Dan PMSDocument27 pagesMateri Iht TMS Dan PMSagusNo ratings yet
- Brochure Neural Navigator EssentialsDocument4 pagesBrochure Neural Navigator EssentialsOana CornutiuNo ratings yet
- Bio PsychologyDocument30 pagesBio PsychologyNEHA JUNEJANo ratings yet
- Walsh 2000 PDFDocument7 pagesWalsh 2000 PDFChrysoula GkaniNo ratings yet
- Biomagnetics - Principles and Applications of Biomagnetic Stimulation and Imaging (PDFDrive)Document364 pagesBiomagnetics - Principles and Applications of Biomagnetic Stimulation and Imaging (PDFDrive)Luís Correia100% (1)
- Brain Imaging Technologies and Their Applications in NeuroscienceDocument45 pagesBrain Imaging Technologies and Their Applications in NeuroscienceNeea AvrielNo ratings yet
- Ehe 07Document286 pagesEhe 07api-348272721No ratings yet
- Graded Motor Imagery Modifies Movement Pain, Cortical Excitability and Sensorimotor Function in Complex Regional Pain SyndromeDocument14 pagesGraded Motor Imagery Modifies Movement Pain, Cortical Excitability and Sensorimotor Function in Complex Regional Pain Syndromeseroxat2005No ratings yet
- P12x V10-V14.01122Document85 pagesP12x V10-V14.01122War WarNo ratings yet
- Cognitive Impairment Along The Course of Depression: Non-Pharmacological Treatment OptionsDocument11 pagesCognitive Impairment Along The Course of Depression: Non-Pharmacological Treatment OptionsCarolina PradoNo ratings yet
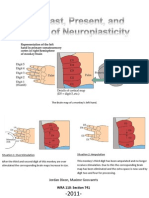
- The Past, Present and Future of NeuroplasticityDocument13 pagesThe Past, Present and Future of NeuroplasticityMaxime Goovaerts100% (3)
- Interview QuestionsDocument5 pagesInterview QuestionsraprapNo ratings yet
- 1-Tecnica y Generalidaddes TMS-EEG 2Document466 pages1-Tecnica y Generalidaddes TMS-EEG 2Lucía del ValleNo ratings yet
- Journal Pre-Proof: CortexDocument34 pagesJournal Pre-Proof: CortexReuben BrownNo ratings yet
- Biological Perspective On Personality: Course: Personality Theories (Psy 711) Lecturer: DR AyodejiDocument24 pagesBiological Perspective On Personality: Course: Personality Theories (Psy 711) Lecturer: DR AyodejiIkponmwosa EseosaNo ratings yet
- Music and Brain Circuitry Strategies For StrengtheningDocument10 pagesMusic and Brain Circuitry Strategies For Strengtheningbiahonda15No ratings yet
- New Treatments in NeurorehabilitationDocument10 pagesNew Treatments in NeurorehabilitationRazvanRotaruNo ratings yet
- 002 Repetitive Transcranial Magentic Stimulation RTMS For Depression PDFDocument114 pages002 Repetitive Transcranial Magentic Stimulation RTMS For Depression PDFISCRR100% (1)
- Better BalanceDocument13 pagesBetter BalanceMiriam NovoNo ratings yet
- Study: Power Changes How The Brain Responds To OthersDocument9 pagesStudy: Power Changes How The Brain Responds To OthersPatricia DillonNo ratings yet
- Boissonnault2021 Article BrainStimulationInAttentionDefDocument9 pagesBoissonnault2021 Article BrainStimulationInAttentionDef奚浩然No ratings yet
- AS Aplicaciones DE LA Estimulación Magnética Transcraneal EN PsiquiatríaDocument10 pagesAS Aplicaciones DE LA Estimulación Magnética Transcraneal EN PsiquiatríaEsmeralda RodriguezNo ratings yet
- Neuromodulation by Means of Electrical Brain Stimulation: AbstractsDocument5 pagesNeuromodulation by Means of Electrical Brain Stimulation: AbstractsmoiNo ratings yet
- INTRODUCCIÓN - (Springer Handbooks) Bruno Siciliano, Oussama Khatib (Eds.) - Springer Handbook of Robotics (2016, Springer)Document5 pagesINTRODUCCIÓN - (Springer Handbooks) Bruno Siciliano, Oussama Khatib (Eds.) - Springer Handbook of Robotics (2016, Springer)Pepe ViyuelaNo ratings yet
- Translating PICO Into Search StrategyDocument29 pagesTranslating PICO Into Search StrategyAhmad SyaukatNo ratings yet
- Paul Allen@roehampton Ac UkDocument22 pagesPaul Allen@roehampton Ac UkGetha ManuabaNo ratings yet