
ATP Synthesis - Biophysics
ATP Synthesis - Biophysics
You might also like
- Science: Modified Strategic Intervention MaterialsDocument28 pagesScience: Modified Strategic Intervention MaterialsSanJoseHS100% (2)
- HW 1Document11 pagesHW 1sajajNo ratings yet
- Plant-Physiology by TAIZ and ZEIGER PDFDocument2 pagesPlant-Physiology by TAIZ and ZEIGER PDFshxxxNo ratings yet
- 8.OxidativePhosphorylation CompleteDocument40 pages8.OxidativePhosphorylation Completeazyhuang77No ratings yet
- Microsoft PowerPoint - Mitocondria - ETC & Oxydative PhosphotylationDocument21 pagesMicrosoft PowerPoint - Mitocondria - ETC & Oxydative PhosphotylationnursurayaeffendyNo ratings yet
- Pages From Lange Bio FlashcardDocument4 pagesPages From Lange Bio Flashcardsepanta2002No ratings yet
- 16.2.3 Oxidative Phosphorylation: Cellular RespirationDocument47 pages16.2.3 Oxidative Phosphorylation: Cellular Respirationeryna sofeaNo ratings yet
- Etc and OxphosDocument54 pagesEtc and Oxphoswajeehgul400No ratings yet
- Electron Transport ChainDocument37 pagesElectron Transport ChainFan SmithNo ratings yet
- Respiration DiagramDocument3 pagesRespiration Diagramapi-521781723No ratings yet
- 1BMS1153 BL1-Energy MetabolismDocument35 pages1BMS1153 BL1-Energy Metabolismshakila786No ratings yet
- 15 OxPhos 3Document10 pages15 OxPhos 3Haze MNo ratings yet
- 0 CHPT 5 - Photosynthesis and Cellular RespirationDocument76 pages0 CHPT 5 - Photosynthesis and Cellular RespirationshinymaterialsNo ratings yet
- Lec 17 (Ch. 14) Oxidative PhosphDocument40 pagesLec 17 (Ch. 14) Oxidative PhosphRamy El-HadadNo ratings yet
- Oxidative PhosphorylationDocument28 pagesOxidative PhosphorylationAnbuNo ratings yet
- FTNS-3101-Oxidative PhosphorylationDocument4 pagesFTNS-3101-Oxidative PhosphorylationMd. Muhaiminul IslamNo ratings yet
- ETC and Oxidative Phosphorylation 2021 NotesDocument9 pagesETC and Oxidative Phosphorylation 2021 NotesmunzleenkashifNo ratings yet
- Electron Transport ChainDocument40 pagesElectron Transport ChainUtkarsh SharmaNo ratings yet
- Bioemergetics Cellular RespDocument59 pagesBioemergetics Cellular RespHassan Abib BasalNo ratings yet
- Electron Transport Chain PDFDocument29 pagesElectron Transport Chain PDFGuru P M100% (2)
- Electron Transport Chain - DETAILED Part 1 AtfDocument6 pagesElectron Transport Chain - DETAILED Part 1 AtfSananNo ratings yet
- Edited Activity 1 8 5Document4 pagesEdited Activity 1 8 5Karl Mathew PajarillagaNo ratings yet
- Photosynthesis March - AprilDocument40 pagesPhotosynthesis March - AprilIbrahim TurayNo ratings yet
- Lecture 03 PDFDocument4 pagesLecture 03 PDFMuhammad TariqNo ratings yet
- Lecture 03 PDFDocument4 pagesLecture 03 PDFRana AmjadNo ratings yet
- Biologic Oxidation & Fosforilasi OksidatifDocument63 pagesBiologic Oxidation & Fosforilasi OksidatifRizky FebriantiNo ratings yet
- Oxidative Phosphorylation 1Document22 pagesOxidative Phosphorylation 1fatin nadia100% (1)
- Bioenergetics MD PhysiothDocument70 pagesBioenergetics MD PhysioththestaffforpediatricptNo ratings yet
- Biological Oxidation: Zhang Hai-Feng Department of Biochemistry, Basic Medical School of Zhengzhou UniversityDocument102 pagesBiological Oxidation: Zhang Hai-Feng Department of Biochemistry, Basic Medical School of Zhengzhou Universityapi-19916399No ratings yet
- FOTOSINTESISDocument27 pagesFOTOSINTESISSumiatinNo ratings yet
- 2 - Cytochromes PDFDocument51 pages2 - Cytochromes PDFAnonymous ojJ64xSOGNo ratings yet
- XAVI PHOTOSYNTHESIS - Lecture BIOUMUMDocument47 pagesXAVI PHOTOSYNTHESIS - Lecture BIOUMUMPrita YhuniNo ratings yet
- 5-Electron Transport Chain - MDDocument31 pages5-Electron Transport Chain - MDIbrahim BarhamNo ratings yet
- Three Stages of Cellular RespirationDocument8 pagesThree Stages of Cellular Respirationapi-418176886No ratings yet
- Teacher - Cellular-RespirationDocument65 pagesTeacher - Cellular-Respirationecha100% (1)
- Cellular Respiration and PhotosyntesisDocument32 pagesCellular Respiration and PhotosyntesisTifahNo ratings yet
- Electrontransportchain 151027073626 Lva1 App6892Document29 pagesElectrontransportchain 151027073626 Lva1 App6892GobindaSahuNo ratings yet
- 1 Oxidative PhosphorylationDocument10 pages1 Oxidative PhosphorylationRoland ToroNo ratings yet
- ATP Synthase StructureDocument36 pagesATP Synthase Structurekiran kombeNo ratings yet
- 5BBB0223 Mitochondrial Function & Dysfunction: Timothy Pullen With Thanks To: DR Clare Thornton, RVCDocument63 pages5BBB0223 Mitochondrial Function & Dysfunction: Timothy Pullen With Thanks To: DR Clare Thornton, RVCW BNo ratings yet
- 4 4 Electron Transport ChainDocument2 pages4 4 Electron Transport ChainKai JieNo ratings yet
- ChemiosmosisDocument7 pagesChemiosmosisSkenzKenzNo ratings yet
- Ch22 Slides Part1 BlankDocument11 pagesCh22 Slides Part1 BlankVictoria ChenNo ratings yet
- Biochem1 Fall2020 LecNov10 PostedDocument57 pagesBiochem1 Fall2020 LecNov10 PostedalexNo ratings yet
- Oksidasi BiologiDocument62 pagesOksidasi BiologisenadaNo ratings yet
- Plan For Today: Finish Up With Electron Transport Chain and Oxidative PhosphorylationDocument52 pagesPlan For Today: Finish Up With Electron Transport Chain and Oxidative PhosphorylationKalai KrishnamurthyNo ratings yet
- Respiration-4 The ETCDocument21 pagesRespiration-4 The ETCSelome KassayeNo ratings yet
- Cellular Respiration Gulf Coast 2013 1Document14 pagesCellular Respiration Gulf Coast 2013 1hamnababar98No ratings yet
- Cells and Sugars 09 Mitochondria and Ox Phos StudentDocument21 pagesCells and Sugars 09 Mitochondria and Ox Phos StudenttyhbbhhNo ratings yet
- Electron Transport Chain - 1Document7 pagesElectron Transport Chain - 1Manash SarmahNo ratings yet
- 001ai Oxidative Phosphorylation PresentationDocument18 pages001ai Oxidative Phosphorylation Presentationbacteria h87No ratings yet
- Electron Transport and Oxidative Phosphorylation: NADH-ubiquinone OxidoreductaseDocument4 pagesElectron Transport and Oxidative Phosphorylation: NADH-ubiquinone OxidoreductaseAries DocNo ratings yet
- BBC2 K27,28RCDocument42 pagesBBC2 K27,28RCMarieta RitongaNo ratings yet
- Diet and ExerciseDocument53 pagesDiet and ExerciseJessica SnowNo ratings yet
- Biological Oxidation (Electron Transport Chain - Chemiosmosis - Oxidative Phosphorylation - Uncouplers)Document22 pagesBiological Oxidation (Electron Transport Chain - Chemiosmosis - Oxidative Phosphorylation - Uncouplers)Krishia GalonaNo ratings yet
- Cellular RespirationDocument45 pagesCellular RespirationRicardo Cubillos UribeNo ratings yet
- Electron Transport ChainDocument35 pagesElectron Transport Chainjanfaiz102No ratings yet
- 21 - N Fixation PDFDocument7 pages21 - N Fixation PDFshubhamNo ratings yet
- Inhibitors & UncouplersDocument7 pagesInhibitors & UncouplersValentino Dhiyu AsmoroNo ratings yet
- Fosforilasi OksidatifDocument82 pagesFosforilasi OksidatifSanti WilujengNo ratings yet
- Electron Transport SystemDocument58 pagesElectron Transport SystemSantosh KumarNo ratings yet
- Iron Metabolism: From Molecular Mechanisms to Clinical ConsequencesFrom EverandIron Metabolism: From Molecular Mechanisms to Clinical ConsequencesRating: 5 out of 5 stars5/5 (1)
- Multiwave 7000: Simplified Method Development For Microwave Acid DigestionDocument21 pagesMultiwave 7000: Simplified Method Development For Microwave Acid DigestiontonyNo ratings yet
- Plastics: Mohammed Bilal 1 N S 1 9 AT 0 2 7 MMBC VDocument21 pagesPlastics: Mohammed Bilal 1 N S 1 9 AT 0 2 7 MMBC Vlalu mikeyNo ratings yet
- Analysis of A Biorefinery Based On Theobroma Grandiflorum (Copoazu) Fruit PDFDocument12 pagesAnalysis of A Biorefinery Based On Theobroma Grandiflorum (Copoazu) Fruit PDFJulio MejiaNo ratings yet
- Raw Data AnalysisDocument20 pagesRaw Data AnalysisSyukri DaimonNo ratings yet
- Solution Practice Test 1 Class 12 - ChemistryDocument6 pagesSolution Practice Test 1 Class 12 - Chemistry39 Yogendra KumarNo ratings yet
- Inorg Chem NotesDocument15 pagesInorg Chem Notesanise santosNo ratings yet
- Author's Accepted Manuscript: Ipomoea BatatasDocument28 pagesAuthor's Accepted Manuscript: Ipomoea BatatasLuqman MileNo ratings yet
- 816-00138-01-Metalworking-Fluids-Selection-Guide DOWDocument38 pages816-00138-01-Metalworking-Fluids-Selection-Guide DOWDidier FARATNo ratings yet
- Silly PuttyDocument3 pagesSilly PuttyPUNDAVELA ANGELICANo ratings yet
- Haloalkanes Haloarenes RAFTAAR Nitesh DevnaniDocument73 pagesHaloalkanes Haloarenes RAFTAAR Nitesh DevnaniTech MilitiaNo ratings yet
- Determination of Acidity & PHDocument3 pagesDetermination of Acidity & PHMonisankar Mula100% (2)
- Acid Number of Petroleum Products by Potentiometric TitrationDocument11 pagesAcid Number of Petroleum Products by Potentiometric TitrationxinyingNo ratings yet
- Review Paper Adenanthera Pavonina PavoninaDocument13 pagesReview Paper Adenanthera Pavonina PavoninaABDUL RUB NAAZNo ratings yet
- WCH11 01 Que 20210304Document24 pagesWCH11 01 Que 20210304윤소리No ratings yet
- Bacterial Physiology and Biochemistry Ivan Kushkevych Full ChapterDocument50 pagesBacterial Physiology and Biochemistry Ivan Kushkevych Full Chapterjean.dunn500100% (8)
- Vinati Organics - Initiating Coverage - 12 Jun 24Document40 pagesVinati Organics - Initiating Coverage - 12 Jun 24amarjeetNo ratings yet
- Analysis of Multiresidue Pesticides in SalmonDocument8 pagesAnalysis of Multiresidue Pesticides in SalmonpeterppsNo ratings yet
- HofmannDocument12 pagesHofmannDavidek VeverkaNo ratings yet
- Obat TitipanDocument52 pagesObat TitipanIsmayantiNo ratings yet
- What Is Solid Waste Management - Sources and Methods of Solid Waste Management - Conserve Energy FutureDocument13 pagesWhat Is Solid Waste Management - Sources and Methods of Solid Waste Management - Conserve Energy FutureBASKAR ENo ratings yet
- NCCSTS CS CompostingDocument9 pagesNCCSTS CS CompostingStricker StikmanNo ratings yet
- Protherm Catalog - 2018Document9 pagesProtherm Catalog - 2018Sunjeev SHARMANo ratings yet
- Biochem LabDocument8 pagesBiochem LabMa Anna Cris LumongsudNo ratings yet
- Catalytic Hydrogenation of Alkynes: As With Alkenes, Addition of Each Pair of H's Is SynDocument6 pagesCatalytic Hydrogenation of Alkynes: As With Alkenes, Addition of Each Pair of H's Is SynVishal SinghNo ratings yet
- Seaweed Liquid Fertilizer (SLF) and Its Role in Agriculture ProductivityDocument11 pagesSeaweed Liquid Fertilizer (SLF) and Its Role in Agriculture ProductivitySamir FellousNo ratings yet
- Organic SheetDocument12 pagesOrganic SheetVengefulNo ratings yet
- 07 Vitamins (Part 1)Document48 pages07 Vitamins (Part 1)NUR ANISAH MOHD ADNANNo ratings yet
- An Efficient, Surfactant Mediated Biginelli Condensation For The One Pot Synthesis of Dihydropyrimidine DerivativesDocument7 pagesAn Efficient, Surfactant Mediated Biginelli Condensation For The One Pot Synthesis of Dihydropyrimidine DerivativesRizka AmaliaNo ratings yet





















































































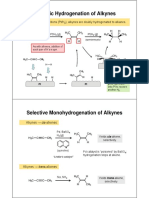




Download as pdf or txt
You might also like
- Science: Modified Strategic Intervention MaterialsDocument28 pagesScience: Modified Strategic Intervention MaterialsSanJoseHS100% (2)
- HW 1Document11 pagesHW 1sajajNo ratings yet
- Plant-Physiology by TAIZ and ZEIGER PDFDocument2 pagesPlant-Physiology by TAIZ and ZEIGER PDFshxxxNo ratings yet
- 8.OxidativePhosphorylation CompleteDocument40 pages8.OxidativePhosphorylation Completeazyhuang77No ratings yet
- Microsoft PowerPoint - Mitocondria - ETC & Oxydative PhosphotylationDocument21 pagesMicrosoft PowerPoint - Mitocondria - ETC & Oxydative PhosphotylationnursurayaeffendyNo ratings yet
- Pages From Lange Bio FlashcardDocument4 pagesPages From Lange Bio Flashcardsepanta2002No ratings yet
- 16.2.3 Oxidative Phosphorylation: Cellular RespirationDocument47 pages16.2.3 Oxidative Phosphorylation: Cellular Respirationeryna sofeaNo ratings yet
- Etc and OxphosDocument54 pagesEtc and Oxphoswajeehgul400No ratings yet
- Electron Transport ChainDocument37 pagesElectron Transport ChainFan SmithNo ratings yet
- Respiration DiagramDocument3 pagesRespiration Diagramapi-521781723No ratings yet
- 1BMS1153 BL1-Energy MetabolismDocument35 pages1BMS1153 BL1-Energy Metabolismshakila786No ratings yet
- 15 OxPhos 3Document10 pages15 OxPhos 3Haze MNo ratings yet
- 0 CHPT 5 - Photosynthesis and Cellular RespirationDocument76 pages0 CHPT 5 - Photosynthesis and Cellular RespirationshinymaterialsNo ratings yet
- Lec 17 (Ch. 14) Oxidative PhosphDocument40 pagesLec 17 (Ch. 14) Oxidative PhosphRamy El-HadadNo ratings yet
- Oxidative PhosphorylationDocument28 pagesOxidative PhosphorylationAnbuNo ratings yet
- FTNS-3101-Oxidative PhosphorylationDocument4 pagesFTNS-3101-Oxidative PhosphorylationMd. Muhaiminul IslamNo ratings yet
- ETC and Oxidative Phosphorylation 2021 NotesDocument9 pagesETC and Oxidative Phosphorylation 2021 NotesmunzleenkashifNo ratings yet
- Electron Transport ChainDocument40 pagesElectron Transport ChainUtkarsh SharmaNo ratings yet
- Bioemergetics Cellular RespDocument59 pagesBioemergetics Cellular RespHassan Abib BasalNo ratings yet
- Electron Transport Chain PDFDocument29 pagesElectron Transport Chain PDFGuru P M100% (2)
- Electron Transport Chain - DETAILED Part 1 AtfDocument6 pagesElectron Transport Chain - DETAILED Part 1 AtfSananNo ratings yet
- Edited Activity 1 8 5Document4 pagesEdited Activity 1 8 5Karl Mathew PajarillagaNo ratings yet
- Photosynthesis March - AprilDocument40 pagesPhotosynthesis March - AprilIbrahim TurayNo ratings yet
- Lecture 03 PDFDocument4 pagesLecture 03 PDFMuhammad TariqNo ratings yet
- Lecture 03 PDFDocument4 pagesLecture 03 PDFRana AmjadNo ratings yet
- Biologic Oxidation & Fosforilasi OksidatifDocument63 pagesBiologic Oxidation & Fosforilasi OksidatifRizky FebriantiNo ratings yet
- Oxidative Phosphorylation 1Document22 pagesOxidative Phosphorylation 1fatin nadia100% (1)
- Bioenergetics MD PhysiothDocument70 pagesBioenergetics MD PhysioththestaffforpediatricptNo ratings yet
- Biological Oxidation: Zhang Hai-Feng Department of Biochemistry, Basic Medical School of Zhengzhou UniversityDocument102 pagesBiological Oxidation: Zhang Hai-Feng Department of Biochemistry, Basic Medical School of Zhengzhou Universityapi-19916399No ratings yet
- FOTOSINTESISDocument27 pagesFOTOSINTESISSumiatinNo ratings yet
- 2 - Cytochromes PDFDocument51 pages2 - Cytochromes PDFAnonymous ojJ64xSOGNo ratings yet
- XAVI PHOTOSYNTHESIS - Lecture BIOUMUMDocument47 pagesXAVI PHOTOSYNTHESIS - Lecture BIOUMUMPrita YhuniNo ratings yet
- 5-Electron Transport Chain - MDDocument31 pages5-Electron Transport Chain - MDIbrahim BarhamNo ratings yet
- Three Stages of Cellular RespirationDocument8 pagesThree Stages of Cellular Respirationapi-418176886No ratings yet
- Teacher - Cellular-RespirationDocument65 pagesTeacher - Cellular-Respirationecha100% (1)
- Cellular Respiration and PhotosyntesisDocument32 pagesCellular Respiration and PhotosyntesisTifahNo ratings yet
- Electrontransportchain 151027073626 Lva1 App6892Document29 pagesElectrontransportchain 151027073626 Lva1 App6892GobindaSahuNo ratings yet
- 1 Oxidative PhosphorylationDocument10 pages1 Oxidative PhosphorylationRoland ToroNo ratings yet
- ATP Synthase StructureDocument36 pagesATP Synthase Structurekiran kombeNo ratings yet
- 5BBB0223 Mitochondrial Function & Dysfunction: Timothy Pullen With Thanks To: DR Clare Thornton, RVCDocument63 pages5BBB0223 Mitochondrial Function & Dysfunction: Timothy Pullen With Thanks To: DR Clare Thornton, RVCW BNo ratings yet
- 4 4 Electron Transport ChainDocument2 pages4 4 Electron Transport ChainKai JieNo ratings yet
- ChemiosmosisDocument7 pagesChemiosmosisSkenzKenzNo ratings yet
- Ch22 Slides Part1 BlankDocument11 pagesCh22 Slides Part1 BlankVictoria ChenNo ratings yet
- Biochem1 Fall2020 LecNov10 PostedDocument57 pagesBiochem1 Fall2020 LecNov10 PostedalexNo ratings yet
- Oksidasi BiologiDocument62 pagesOksidasi BiologisenadaNo ratings yet
- Plan For Today: Finish Up With Electron Transport Chain and Oxidative PhosphorylationDocument52 pagesPlan For Today: Finish Up With Electron Transport Chain and Oxidative PhosphorylationKalai KrishnamurthyNo ratings yet
- Respiration-4 The ETCDocument21 pagesRespiration-4 The ETCSelome KassayeNo ratings yet
- Cellular Respiration Gulf Coast 2013 1Document14 pagesCellular Respiration Gulf Coast 2013 1hamnababar98No ratings yet
- Cells and Sugars 09 Mitochondria and Ox Phos StudentDocument21 pagesCells and Sugars 09 Mitochondria and Ox Phos StudenttyhbbhhNo ratings yet
- Electron Transport Chain - 1Document7 pagesElectron Transport Chain - 1Manash SarmahNo ratings yet
- 001ai Oxidative Phosphorylation PresentationDocument18 pages001ai Oxidative Phosphorylation Presentationbacteria h87No ratings yet
- Electron Transport and Oxidative Phosphorylation: NADH-ubiquinone OxidoreductaseDocument4 pagesElectron Transport and Oxidative Phosphorylation: NADH-ubiquinone OxidoreductaseAries DocNo ratings yet
- BBC2 K27,28RCDocument42 pagesBBC2 K27,28RCMarieta RitongaNo ratings yet
- Diet and ExerciseDocument53 pagesDiet and ExerciseJessica SnowNo ratings yet
- Biological Oxidation (Electron Transport Chain - Chemiosmosis - Oxidative Phosphorylation - Uncouplers)Document22 pagesBiological Oxidation (Electron Transport Chain - Chemiosmosis - Oxidative Phosphorylation - Uncouplers)Krishia GalonaNo ratings yet
- Cellular RespirationDocument45 pagesCellular RespirationRicardo Cubillos UribeNo ratings yet
- Electron Transport ChainDocument35 pagesElectron Transport Chainjanfaiz102No ratings yet
- 21 - N Fixation PDFDocument7 pages21 - N Fixation PDFshubhamNo ratings yet
- Inhibitors & UncouplersDocument7 pagesInhibitors & UncouplersValentino Dhiyu AsmoroNo ratings yet
- Fosforilasi OksidatifDocument82 pagesFosforilasi OksidatifSanti WilujengNo ratings yet
- Electron Transport SystemDocument58 pagesElectron Transport SystemSantosh KumarNo ratings yet
- Iron Metabolism: From Molecular Mechanisms to Clinical ConsequencesFrom EverandIron Metabolism: From Molecular Mechanisms to Clinical ConsequencesRating: 5 out of 5 stars5/5 (1)
- Multiwave 7000: Simplified Method Development For Microwave Acid DigestionDocument21 pagesMultiwave 7000: Simplified Method Development For Microwave Acid DigestiontonyNo ratings yet
- Plastics: Mohammed Bilal 1 N S 1 9 AT 0 2 7 MMBC VDocument21 pagesPlastics: Mohammed Bilal 1 N S 1 9 AT 0 2 7 MMBC Vlalu mikeyNo ratings yet
- Analysis of A Biorefinery Based On Theobroma Grandiflorum (Copoazu) Fruit PDFDocument12 pagesAnalysis of A Biorefinery Based On Theobroma Grandiflorum (Copoazu) Fruit PDFJulio MejiaNo ratings yet
- Raw Data AnalysisDocument20 pagesRaw Data AnalysisSyukri DaimonNo ratings yet
- Solution Practice Test 1 Class 12 - ChemistryDocument6 pagesSolution Practice Test 1 Class 12 - Chemistry39 Yogendra KumarNo ratings yet
- Inorg Chem NotesDocument15 pagesInorg Chem Notesanise santosNo ratings yet
- Author's Accepted Manuscript: Ipomoea BatatasDocument28 pagesAuthor's Accepted Manuscript: Ipomoea BatatasLuqman MileNo ratings yet
- 816-00138-01-Metalworking-Fluids-Selection-Guide DOWDocument38 pages816-00138-01-Metalworking-Fluids-Selection-Guide DOWDidier FARATNo ratings yet
- Silly PuttyDocument3 pagesSilly PuttyPUNDAVELA ANGELICANo ratings yet
- Haloalkanes Haloarenes RAFTAAR Nitesh DevnaniDocument73 pagesHaloalkanes Haloarenes RAFTAAR Nitesh DevnaniTech MilitiaNo ratings yet
- Determination of Acidity & PHDocument3 pagesDetermination of Acidity & PHMonisankar Mula100% (2)
- Acid Number of Petroleum Products by Potentiometric TitrationDocument11 pagesAcid Number of Petroleum Products by Potentiometric TitrationxinyingNo ratings yet
- Review Paper Adenanthera Pavonina PavoninaDocument13 pagesReview Paper Adenanthera Pavonina PavoninaABDUL RUB NAAZNo ratings yet
- WCH11 01 Que 20210304Document24 pagesWCH11 01 Que 20210304윤소리No ratings yet
- Bacterial Physiology and Biochemistry Ivan Kushkevych Full ChapterDocument50 pagesBacterial Physiology and Biochemistry Ivan Kushkevych Full Chapterjean.dunn500100% (8)
- Vinati Organics - Initiating Coverage - 12 Jun 24Document40 pagesVinati Organics - Initiating Coverage - 12 Jun 24amarjeetNo ratings yet
- Analysis of Multiresidue Pesticides in SalmonDocument8 pagesAnalysis of Multiresidue Pesticides in SalmonpeterppsNo ratings yet
- HofmannDocument12 pagesHofmannDavidek VeverkaNo ratings yet
- Obat TitipanDocument52 pagesObat TitipanIsmayantiNo ratings yet
- What Is Solid Waste Management - Sources and Methods of Solid Waste Management - Conserve Energy FutureDocument13 pagesWhat Is Solid Waste Management - Sources and Methods of Solid Waste Management - Conserve Energy FutureBASKAR ENo ratings yet
- NCCSTS CS CompostingDocument9 pagesNCCSTS CS CompostingStricker StikmanNo ratings yet
- Protherm Catalog - 2018Document9 pagesProtherm Catalog - 2018Sunjeev SHARMANo ratings yet
- Biochem LabDocument8 pagesBiochem LabMa Anna Cris LumongsudNo ratings yet
- Catalytic Hydrogenation of Alkynes: As With Alkenes, Addition of Each Pair of H's Is SynDocument6 pagesCatalytic Hydrogenation of Alkynes: As With Alkenes, Addition of Each Pair of H's Is SynVishal SinghNo ratings yet
- Seaweed Liquid Fertilizer (SLF) and Its Role in Agriculture ProductivityDocument11 pagesSeaweed Liquid Fertilizer (SLF) and Its Role in Agriculture ProductivitySamir FellousNo ratings yet
- Organic SheetDocument12 pagesOrganic SheetVengefulNo ratings yet
- 07 Vitamins (Part 1)Document48 pages07 Vitamins (Part 1)NUR ANISAH MOHD ADNANNo ratings yet
- An Efficient, Surfactant Mediated Biginelli Condensation For The One Pot Synthesis of Dihydropyrimidine DerivativesDocument7 pagesAn Efficient, Surfactant Mediated Biginelli Condensation For The One Pot Synthesis of Dihydropyrimidine DerivativesRizka AmaliaNo ratings yet