
Principles of Diabetes Mellitus - Ch3Final
Principles of Diabetes Mellitus - Ch3Final
You might also like
- Active Reading: Answer KeyDocument3 pagesActive Reading: Answer KeyJonas Montero100% (2)
- Fisiologi of PankreasDocument9 pagesFisiologi of PankreasSunardi BahriNo ratings yet
- Physiology, Endocrine, Pancreas - StatPearls - NCBI BookshelfDocument5 pagesPhysiology, Endocrine, Pancreas - StatPearls - NCBI Bookshelfdrpaez27No ratings yet
- InsulinDocument50 pagesInsulinTandin SonamNo ratings yet
- Insulin 2 PDFDocument15 pagesInsulin 2 PDFAnonymous K9tmd7T0PzNo ratings yet
- 3 Types of Organism As A ProductDocument2 pages3 Types of Organism As A ProductShaistha FriasNo ratings yet
- Insulinoma in Dogs: A Review: Normal Glucose HomeostasisDocument13 pagesInsulinoma in Dogs: A Review: Normal Glucose HomeostasisBrisbany AlcibarNo ratings yet
- Nutrient Regulation of InsulinDocument16 pagesNutrient Regulation of InsulinNickolasMarllesNo ratings yet
- Mechanism Controlling Pancreatic Islet Cell Function in Insulin SecretionDocument17 pagesMechanism Controlling Pancreatic Islet Cell Function in Insulin SecretionDahiana BergesNo ratings yet
- Physiology of Glucose Homeostasis and Insulin Therapy in Type 1 and Type 2 Diabetes (2012)Document15 pagesPhysiology of Glucose Homeostasis and Insulin Therapy in Type 1 and Type 2 Diabetes (2012)drheayNo ratings yet
- 1 s2.0 S0022282816302176 MainDocument85 pages1 s2.0 S0022282816302176 MainNia Rukman100% (1)
- Regenerative Medicine in Diabetes: General IntroductionDocument10 pagesRegenerative Medicine in Diabetes: General IntroductionAlex PlicNo ratings yet
- Insuline HormoneDocument36 pagesInsuline HormoneF ParikhNo ratings yet
- Noshaba MSDocument3 pagesNoshaba MSKb AliNo ratings yet
- Endo Pancrease ElhDocument27 pagesEndo Pancrease Elhodiodi57No ratings yet
- InsulinDocument66 pagesInsulinbhatiaharryjassiNo ratings yet
- Physio 7Document12 pagesPhysio 7amjadNo ratings yet
- Insulin Bio SynthesisDocument12 pagesInsulin Bio SynthesisdrtpkNo ratings yet
- (OS 202C) 20120102 Pancreatic Islet Physiology (Insulin)Document10 pages(OS 202C) 20120102 Pancreatic Islet Physiology (Insulin)niaasetaNo ratings yet
- Role of Incretins in Pancreas Growth and Development: EditorialDocument3 pagesRole of Incretins in Pancreas Growth and Development: EditorialHarsha Ramakrishna KashyapNo ratings yet
- KersenDocument13 pagesKersenJumasni AdnanNo ratings yet
- Insulin, Glucagon, and Diabetes Mellitus: Insulin Is A Hormone Associated With Energy AbundanceDocument17 pagesInsulin, Glucagon, and Diabetes Mellitus: Insulin Is A Hormone Associated With Energy AbundancedawnNo ratings yet
- Pancreas PancreasDocument18 pagesPancreas PancreasZzeba KhanNo ratings yet
- GH Chap 78 - Insulin, Glucagon and DMDocument16 pagesGH Chap 78 - Insulin, Glucagon and DMShellz2428No ratings yet
- The Role of Parathyroid Hormone-Related Protein (PTHRP) in The Pathophysiology of Diabetes MellitusDocument7 pagesThe Role of Parathyroid Hormone-Related Protein (PTHRP) in The Pathophysiology of Diabetes Mellitusfachrudin arrozaqNo ratings yet
- Endocrin E Pancreas: DR - Sabri.S.EltaybeDocument43 pagesEndocrin E Pancreas: DR - Sabri.S.EltaybehamidNo ratings yet
- Chapter 60 - Insulin, Oral Hypoglycaemic Agents, and The Pharmacology of The Endocrine PancreasDocument17 pagesChapter 60 - Insulin, Oral Hypoglycaemic Agents, and The Pharmacology of The Endocrine PancreasGiliana P. García AcevedoNo ratings yet
- Insulin EndotextDocument37 pagesInsulin EndotextHendra HamzahNo ratings yet
- Endocrine System - MCAT Biology 5Document9 pagesEndocrine System - MCAT Biology 5Saran p sNo ratings yet
- Pancreas Review Nature 2018Document8 pagesPancreas Review Nature 2018Geovanny ImuesNo ratings yet
- Insulin and The Oral HypoglycemicsDocument9 pagesInsulin and The Oral HypoglycemicsHasibNo ratings yet
- The Integrative Biology of Type 2 DiabetesDocument10 pagesThe Integrative Biology of Type 2 DiabetesAdrian GhiţăNo ratings yet
- Suppression of Insulin Production and Secretion by A Decretin HormoneDocument12 pagesSuppression of Insulin Production and Secretion by A Decretin HormoneMemoo MirizioNo ratings yet
- Pancreatic Regulation of Glucose HomeostasisDocument20 pagesPancreatic Regulation of Glucose Homeostasiseki_herawatiNo ratings yet
- Liu 2014Document8 pagesLiu 201423501122No ratings yet
- Insulin Secretion: 1. Pancreatic Beta CellsDocument6 pagesInsulin Secretion: 1. Pancreatic Beta CellsAlicia Tormos EsteveNo ratings yet
- Direct Effect of Cholesterol On Insulin Secretion A Novel Mechanism For Pancreatic ?-Cell DysfunctionDocument11 pagesDirect Effect of Cholesterol On Insulin Secretion A Novel Mechanism For Pancreatic ?-Cell DysfunctionAsmat BurhanNo ratings yet
- Đề sinh hóa vấn đápDocument2 pagesĐề sinh hóa vấn đápNghĩa Bùi Lê QuangNo ratings yet
- Proof: Roles of Amylin in Satiation, Adiposity and Brain DevelopmentDocument11 pagesProof: Roles of Amylin in Satiation, Adiposity and Brain DevelopmentBruno Melo-FerreiraNo ratings yet
- Lept in 012Document7 pagesLept in 012Carolina De Lima FerreiraNo ratings yet
- tmp24D8 TMPDocument13 pagestmp24D8 TMPFrontiersNo ratings yet
- GI and Reproductive HormoneDocument74 pagesGI and Reproductive Hormonepranutan739No ratings yet
- Diabetes Spectr 2004 Aronoff 183 90Document8 pagesDiabetes Spectr 2004 Aronoff 183 90Bodea DoruNo ratings yet
- Metabolisme Insulin Dan GlukagonDocument7 pagesMetabolisme Insulin Dan GlukagonDinda saylaNo ratings yet
- Downsi 6 PDFDocument11 pagesDownsi 6 PDFmutimutimutiNo ratings yet
- Hormonal Regulation of MetabolismDocument19 pagesHormonal Regulation of MetabolismnizaNo ratings yet
- Obesity and Insulin Resistance: Barbara B. Kahn and Jeffrey S. FlierDocument9 pagesObesity and Insulin Resistance: Barbara B. Kahn and Jeffrey S. FlierernaataNo ratings yet
- Obesity and Insulin Resistance: Barbara B. Kahn and Jeffrey S. FlierDocument9 pagesObesity and Insulin Resistance: Barbara B. Kahn and Jeffrey S. Fliersapphire_asaphNo ratings yet
- Nutrients 03 00735Document21 pagesNutrients 03 00735Yvett HorváthNo ratings yet
- Diabetic Emergencies in Small AnimalsDocument17 pagesDiabetic Emergencies in Small Animalsdia_dianneNo ratings yet
- Homeostasis and The Regulation of Blood GlucoseDocument7 pagesHomeostasis and The Regulation of Blood GlucoseElgin SammonsNo ratings yet
- DownloadDocument11 pagesDownloadadri90No ratings yet
- Gastrointestinal Physiology - Mosby Physiology Monograph Series (Mosby's Physiology Monograph)Document6 pagesGastrointestinal Physiology - Mosby Physiology Monograph Series (Mosby's Physiology Monograph)leonorsimoesfernandesNo ratings yet
- The Role of Polyunsaturated Fatty Acids N 3 PUFAs On The Pancreatic Cells and Insulin ActionDocument8 pagesThe Role of Polyunsaturated Fatty Acids N 3 PUFAs On The Pancreatic Cells and Insulin ActionHabtamu WondifrawNo ratings yet
- Adiponectin - Journey From An Adipocyte SecretoryDocument9 pagesAdiponectin - Journey From An Adipocyte SecretorygiannidietNo ratings yet
- Aspects Physiologiques Et Métaboliques de Contrôle de L'ingestion AlimentaireDocument8 pagesAspects Physiologiques Et Métaboliques de Contrôle de L'ingestion Alimentairesoussou.dellacheNo ratings yet
- Nature 14190Document9 pagesNature 14190mike22tfNo ratings yet
- Insulin's Mechanism of ActionDocument14 pagesInsulin's Mechanism of ActionKishore ChinnaNo ratings yet
- Diabetes 2001 RonnerDocument10 pagesDiabetes 2001 RonnerGerardo Félix MartínezNo ratings yet
- Insulin ResistanceDocument13 pagesInsulin ResistanceKlinik Hanis0% (1)
- Leptin: Regulation and Clinical ApplicationsFrom EverandLeptin: Regulation and Clinical ApplicationsSam Dagogo-Jack, MDNo ratings yet
- Olimpiada Engleza Arad - 2016 PDFDocument5 pagesOlimpiada Engleza Arad - 2016 PDFAnthony AdamsNo ratings yet
- Rancho Foods Deposits All Cash Receipts Each Wednesday and FridayDocument1 pageRancho Foods Deposits All Cash Receipts Each Wednesday and FridayAmit PandeyNo ratings yet
- Simulated Binar Crossover F Continuous Search Space: Comp SystemsDocument34 pagesSimulated Binar Crossover F Continuous Search Space: Comp SystemsSal Ie EmNo ratings yet
- Garia - Wikipedia, The Free EncyclopediaDocument9 pagesGaria - Wikipedia, The Free EncyclopediaPradip DattaNo ratings yet
- Interim Report - SampleDocument34 pagesInterim Report - SampleRISHAV DUTTANo ratings yet
- Montreal Protocol NotesDocument2 pagesMontreal Protocol NotesSidemen For LifeNo ratings yet
- Nature of BusinessDocument4 pagesNature of BusinessLeslie Ann IgnacioNo ratings yet
- L36GL Data Sheet PDFDocument2 pagesL36GL Data Sheet PDFManuel ValdiviesoNo ratings yet
- LacusCurtius - Strabo's Geography - Book III Chapter 3Document10 pagesLacusCurtius - Strabo's Geography - Book III Chapter 3A Miguel Simão LealNo ratings yet
- MEETING 5 - Describing PeopleDocument6 pagesMEETING 5 - Describing Peoplerizky.ameliaNo ratings yet
- MUET HDocument2 pagesMUET HWONG LUO EE EUNICENo ratings yet
- NCM 205 LabDocument4 pagesNCM 205 LabAlthea Meg MasicampoNo ratings yet
- Synthesis and Characterization of Cadmium Sulphide NanoparticlesDocument17 pagesSynthesis and Characterization of Cadmium Sulphide NanoparticlesHoneylyn IgnacioNo ratings yet
- Practical File Part-1Document136 pagesPractical File Part-1soniya tewatiaNo ratings yet
- MMDocument1 pageMMMark Michael ResuelloNo ratings yet
- Tcoa S A0008208710 1-2563780Document5 pagesTcoa S A0008208710 1-2563780Axel BelzitiNo ratings yet
- RPV 3Document158 pagesRPV 3Melkamu AmusheNo ratings yet
- Cage Batterie PondeuseDocument8 pagesCage Batterie PondeuseSébastien Sylvestre RakotonirianaNo ratings yet
- UntitledDocument108 pagesUntitledSasquarch VeinNo ratings yet
- Contemporary Business 16th Edition Boone Test Bank 1Document29 pagesContemporary Business 16th Edition Boone Test Bank 1mary100% (36)
- Cost ReductionDocument8 pagesCost Reductionmlganesh666100% (3)
- Strategic Management Lecture NotesDocument117 pagesStrategic Management Lecture NotesTinashe Mazorodze100% (2)
- Acharya Nagarjuna University: Examination Application FormDocument2 pagesAcharya Nagarjuna University: Examination Application FormMaragani MuraligangadhararaoNo ratings yet
- Design of Engine Stand For Inline Type Engines. Final Year ProjectDocument71 pagesDesign of Engine Stand For Inline Type Engines. Final Year ProjectTolasa NamomsaNo ratings yet
- PA4600 & PA6000 User Manual - Rev GDocument53 pagesPA4600 & PA6000 User Manual - Rev GjatzireNo ratings yet
- Wago-I/O-System 750: ManualDocument46 pagesWago-I/O-System 750: ManualHaris bayu kurniawanNo ratings yet
- The Financial Statements of Divine Company Are Presented Here DivineDocument1 pageThe Financial Statements of Divine Company Are Presented Here Divineamit raajNo ratings yet
- Li ReportDocument47 pagesLi ReportRizal RizalmanNo ratings yet
- Retroactive Inspection ReportDocument4 pagesRetroactive Inspection Reportdpsy1245No ratings yet






















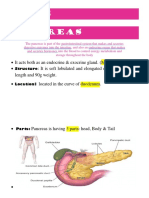



































































Download as pdf or txt
You might also like
- Active Reading: Answer KeyDocument3 pagesActive Reading: Answer KeyJonas Montero100% (2)
- Fisiologi of PankreasDocument9 pagesFisiologi of PankreasSunardi BahriNo ratings yet
- Physiology, Endocrine, Pancreas - StatPearls - NCBI BookshelfDocument5 pagesPhysiology, Endocrine, Pancreas - StatPearls - NCBI Bookshelfdrpaez27No ratings yet
- InsulinDocument50 pagesInsulinTandin SonamNo ratings yet
- Insulin 2 PDFDocument15 pagesInsulin 2 PDFAnonymous K9tmd7T0PzNo ratings yet
- 3 Types of Organism As A ProductDocument2 pages3 Types of Organism As A ProductShaistha FriasNo ratings yet
- Insulinoma in Dogs: A Review: Normal Glucose HomeostasisDocument13 pagesInsulinoma in Dogs: A Review: Normal Glucose HomeostasisBrisbany AlcibarNo ratings yet
- Nutrient Regulation of InsulinDocument16 pagesNutrient Regulation of InsulinNickolasMarllesNo ratings yet
- Mechanism Controlling Pancreatic Islet Cell Function in Insulin SecretionDocument17 pagesMechanism Controlling Pancreatic Islet Cell Function in Insulin SecretionDahiana BergesNo ratings yet
- Physiology of Glucose Homeostasis and Insulin Therapy in Type 1 and Type 2 Diabetes (2012)Document15 pagesPhysiology of Glucose Homeostasis and Insulin Therapy in Type 1 and Type 2 Diabetes (2012)drheayNo ratings yet
- 1 s2.0 S0022282816302176 MainDocument85 pages1 s2.0 S0022282816302176 MainNia Rukman100% (1)
- Regenerative Medicine in Diabetes: General IntroductionDocument10 pagesRegenerative Medicine in Diabetes: General IntroductionAlex PlicNo ratings yet
- Insuline HormoneDocument36 pagesInsuline HormoneF ParikhNo ratings yet
- Noshaba MSDocument3 pagesNoshaba MSKb AliNo ratings yet
- Endo Pancrease ElhDocument27 pagesEndo Pancrease Elhodiodi57No ratings yet
- InsulinDocument66 pagesInsulinbhatiaharryjassiNo ratings yet
- Physio 7Document12 pagesPhysio 7amjadNo ratings yet
- Insulin Bio SynthesisDocument12 pagesInsulin Bio SynthesisdrtpkNo ratings yet
- (OS 202C) 20120102 Pancreatic Islet Physiology (Insulin)Document10 pages(OS 202C) 20120102 Pancreatic Islet Physiology (Insulin)niaasetaNo ratings yet
- Role of Incretins in Pancreas Growth and Development: EditorialDocument3 pagesRole of Incretins in Pancreas Growth and Development: EditorialHarsha Ramakrishna KashyapNo ratings yet
- KersenDocument13 pagesKersenJumasni AdnanNo ratings yet
- Insulin, Glucagon, and Diabetes Mellitus: Insulin Is A Hormone Associated With Energy AbundanceDocument17 pagesInsulin, Glucagon, and Diabetes Mellitus: Insulin Is A Hormone Associated With Energy AbundancedawnNo ratings yet
- Pancreas PancreasDocument18 pagesPancreas PancreasZzeba KhanNo ratings yet
- GH Chap 78 - Insulin, Glucagon and DMDocument16 pagesGH Chap 78 - Insulin, Glucagon and DMShellz2428No ratings yet
- The Role of Parathyroid Hormone-Related Protein (PTHRP) in The Pathophysiology of Diabetes MellitusDocument7 pagesThe Role of Parathyroid Hormone-Related Protein (PTHRP) in The Pathophysiology of Diabetes Mellitusfachrudin arrozaqNo ratings yet
- Endocrin E Pancreas: DR - Sabri.S.EltaybeDocument43 pagesEndocrin E Pancreas: DR - Sabri.S.EltaybehamidNo ratings yet
- Chapter 60 - Insulin, Oral Hypoglycaemic Agents, and The Pharmacology of The Endocrine PancreasDocument17 pagesChapter 60 - Insulin, Oral Hypoglycaemic Agents, and The Pharmacology of The Endocrine PancreasGiliana P. García AcevedoNo ratings yet
- Insulin EndotextDocument37 pagesInsulin EndotextHendra HamzahNo ratings yet
- Endocrine System - MCAT Biology 5Document9 pagesEndocrine System - MCAT Biology 5Saran p sNo ratings yet
- Pancreas Review Nature 2018Document8 pagesPancreas Review Nature 2018Geovanny ImuesNo ratings yet
- Insulin and The Oral HypoglycemicsDocument9 pagesInsulin and The Oral HypoglycemicsHasibNo ratings yet
- The Integrative Biology of Type 2 DiabetesDocument10 pagesThe Integrative Biology of Type 2 DiabetesAdrian GhiţăNo ratings yet
- Suppression of Insulin Production and Secretion by A Decretin HormoneDocument12 pagesSuppression of Insulin Production and Secretion by A Decretin HormoneMemoo MirizioNo ratings yet
- Pancreatic Regulation of Glucose HomeostasisDocument20 pagesPancreatic Regulation of Glucose Homeostasiseki_herawatiNo ratings yet
- Liu 2014Document8 pagesLiu 201423501122No ratings yet
- Insulin Secretion: 1. Pancreatic Beta CellsDocument6 pagesInsulin Secretion: 1. Pancreatic Beta CellsAlicia Tormos EsteveNo ratings yet
- Direct Effect of Cholesterol On Insulin Secretion A Novel Mechanism For Pancreatic ?-Cell DysfunctionDocument11 pagesDirect Effect of Cholesterol On Insulin Secretion A Novel Mechanism For Pancreatic ?-Cell DysfunctionAsmat BurhanNo ratings yet
- Đề sinh hóa vấn đápDocument2 pagesĐề sinh hóa vấn đápNghĩa Bùi Lê QuangNo ratings yet
- Proof: Roles of Amylin in Satiation, Adiposity and Brain DevelopmentDocument11 pagesProof: Roles of Amylin in Satiation, Adiposity and Brain DevelopmentBruno Melo-FerreiraNo ratings yet
- Lept in 012Document7 pagesLept in 012Carolina De Lima FerreiraNo ratings yet
- tmp24D8 TMPDocument13 pagestmp24D8 TMPFrontiersNo ratings yet
- GI and Reproductive HormoneDocument74 pagesGI and Reproductive Hormonepranutan739No ratings yet
- Diabetes Spectr 2004 Aronoff 183 90Document8 pagesDiabetes Spectr 2004 Aronoff 183 90Bodea DoruNo ratings yet
- Metabolisme Insulin Dan GlukagonDocument7 pagesMetabolisme Insulin Dan GlukagonDinda saylaNo ratings yet
- Downsi 6 PDFDocument11 pagesDownsi 6 PDFmutimutimutiNo ratings yet
- Hormonal Regulation of MetabolismDocument19 pagesHormonal Regulation of MetabolismnizaNo ratings yet
- Obesity and Insulin Resistance: Barbara B. Kahn and Jeffrey S. FlierDocument9 pagesObesity and Insulin Resistance: Barbara B. Kahn and Jeffrey S. FlierernaataNo ratings yet
- Obesity and Insulin Resistance: Barbara B. Kahn and Jeffrey S. FlierDocument9 pagesObesity and Insulin Resistance: Barbara B. Kahn and Jeffrey S. Fliersapphire_asaphNo ratings yet
- Nutrients 03 00735Document21 pagesNutrients 03 00735Yvett HorváthNo ratings yet
- Diabetic Emergencies in Small AnimalsDocument17 pagesDiabetic Emergencies in Small Animalsdia_dianneNo ratings yet
- Homeostasis and The Regulation of Blood GlucoseDocument7 pagesHomeostasis and The Regulation of Blood GlucoseElgin SammonsNo ratings yet
- DownloadDocument11 pagesDownloadadri90No ratings yet
- Gastrointestinal Physiology - Mosby Physiology Monograph Series (Mosby's Physiology Monograph)Document6 pagesGastrointestinal Physiology - Mosby Physiology Monograph Series (Mosby's Physiology Monograph)leonorsimoesfernandesNo ratings yet
- The Role of Polyunsaturated Fatty Acids N 3 PUFAs On The Pancreatic Cells and Insulin ActionDocument8 pagesThe Role of Polyunsaturated Fatty Acids N 3 PUFAs On The Pancreatic Cells and Insulin ActionHabtamu WondifrawNo ratings yet
- Adiponectin - Journey From An Adipocyte SecretoryDocument9 pagesAdiponectin - Journey From An Adipocyte SecretorygiannidietNo ratings yet
- Aspects Physiologiques Et Métaboliques de Contrôle de L'ingestion AlimentaireDocument8 pagesAspects Physiologiques Et Métaboliques de Contrôle de L'ingestion Alimentairesoussou.dellacheNo ratings yet
- Nature 14190Document9 pagesNature 14190mike22tfNo ratings yet
- Insulin's Mechanism of ActionDocument14 pagesInsulin's Mechanism of ActionKishore ChinnaNo ratings yet
- Diabetes 2001 RonnerDocument10 pagesDiabetes 2001 RonnerGerardo Félix MartínezNo ratings yet
- Insulin ResistanceDocument13 pagesInsulin ResistanceKlinik Hanis0% (1)
- Leptin: Regulation and Clinical ApplicationsFrom EverandLeptin: Regulation and Clinical ApplicationsSam Dagogo-Jack, MDNo ratings yet
- Olimpiada Engleza Arad - 2016 PDFDocument5 pagesOlimpiada Engleza Arad - 2016 PDFAnthony AdamsNo ratings yet
- Rancho Foods Deposits All Cash Receipts Each Wednesday and FridayDocument1 pageRancho Foods Deposits All Cash Receipts Each Wednesday and FridayAmit PandeyNo ratings yet
- Simulated Binar Crossover F Continuous Search Space: Comp SystemsDocument34 pagesSimulated Binar Crossover F Continuous Search Space: Comp SystemsSal Ie EmNo ratings yet
- Garia - Wikipedia, The Free EncyclopediaDocument9 pagesGaria - Wikipedia, The Free EncyclopediaPradip DattaNo ratings yet
- Interim Report - SampleDocument34 pagesInterim Report - SampleRISHAV DUTTANo ratings yet
- Montreal Protocol NotesDocument2 pagesMontreal Protocol NotesSidemen For LifeNo ratings yet
- Nature of BusinessDocument4 pagesNature of BusinessLeslie Ann IgnacioNo ratings yet
- L36GL Data Sheet PDFDocument2 pagesL36GL Data Sheet PDFManuel ValdiviesoNo ratings yet
- LacusCurtius - Strabo's Geography - Book III Chapter 3Document10 pagesLacusCurtius - Strabo's Geography - Book III Chapter 3A Miguel Simão LealNo ratings yet
- MEETING 5 - Describing PeopleDocument6 pagesMEETING 5 - Describing Peoplerizky.ameliaNo ratings yet
- MUET HDocument2 pagesMUET HWONG LUO EE EUNICENo ratings yet
- NCM 205 LabDocument4 pagesNCM 205 LabAlthea Meg MasicampoNo ratings yet
- Synthesis and Characterization of Cadmium Sulphide NanoparticlesDocument17 pagesSynthesis and Characterization of Cadmium Sulphide NanoparticlesHoneylyn IgnacioNo ratings yet
- Practical File Part-1Document136 pagesPractical File Part-1soniya tewatiaNo ratings yet
- MMDocument1 pageMMMark Michael ResuelloNo ratings yet
- Tcoa S A0008208710 1-2563780Document5 pagesTcoa S A0008208710 1-2563780Axel BelzitiNo ratings yet
- RPV 3Document158 pagesRPV 3Melkamu AmusheNo ratings yet
- Cage Batterie PondeuseDocument8 pagesCage Batterie PondeuseSébastien Sylvestre RakotonirianaNo ratings yet
- UntitledDocument108 pagesUntitledSasquarch VeinNo ratings yet
- Contemporary Business 16th Edition Boone Test Bank 1Document29 pagesContemporary Business 16th Edition Boone Test Bank 1mary100% (36)
- Cost ReductionDocument8 pagesCost Reductionmlganesh666100% (3)
- Strategic Management Lecture NotesDocument117 pagesStrategic Management Lecture NotesTinashe Mazorodze100% (2)
- Acharya Nagarjuna University: Examination Application FormDocument2 pagesAcharya Nagarjuna University: Examination Application FormMaragani MuraligangadhararaoNo ratings yet
- Design of Engine Stand For Inline Type Engines. Final Year ProjectDocument71 pagesDesign of Engine Stand For Inline Type Engines. Final Year ProjectTolasa NamomsaNo ratings yet
- PA4600 & PA6000 User Manual - Rev GDocument53 pagesPA4600 & PA6000 User Manual - Rev GjatzireNo ratings yet
- Wago-I/O-System 750: ManualDocument46 pagesWago-I/O-System 750: ManualHaris bayu kurniawanNo ratings yet
- The Financial Statements of Divine Company Are Presented Here DivineDocument1 pageThe Financial Statements of Divine Company Are Presented Here Divineamit raajNo ratings yet
- Li ReportDocument47 pagesLi ReportRizal RizalmanNo ratings yet
- Retroactive Inspection ReportDocument4 pagesRetroactive Inspection Reportdpsy1245No ratings yet