Download as pdf or txt
You might also like
- CMAR 35175 The Orphan G Protein Receptor Gpr55 As An Emerging Target in 062813Document9 pagesCMAR 35175 The Orphan G Protein Receptor Gpr55 As An Emerging Target in 062813umesh123patilNo ratings yet
- Functional Diversification of Cell Signaling by GPCR LocalizationDocument11 pagesFunctional Diversification of Cell Signaling by GPCR LocalizationTibor SzénásiNo ratings yet
- Gpcrs and CancervistaDocument45 pagesGpcrs and CancervistapriyaaNo ratings yet
- Mini GDocument8 pagesMini GOlivia LuciannoNo ratings yet
- JVPT-2012 Vol11Document108 pagesJVPT-2012 Vol11AlphaRaj Mekapogu100% (1)
- Proteína G en La VisiónDocument16 pagesProteína G en La VisiónJohan PinzonNo ratings yet
- Superfamily of G-Protein Coupled Receptors (GPCRS) - Extraordinary and Outstanding Success of EvolutionDocument13 pagesSuperfamily of G-Protein Coupled Receptors (GPCRS) - Extraordinary and Outstanding Success of EvolutionSELNo ratings yet
- My Journal 2Document11 pagesMy Journal 2FelixNo ratings yet
- Physrev 00021 2020Document51 pagesPhysrev 00021 2020Dani MurilloNo ratings yet
- Interaction of Glucocorticoid Receptor Isoforms With TranscriptionDocument10 pagesInteraction of Glucocorticoid Receptor Isoforms With TranscriptionSyifa SariNo ratings yet
- Orphan Receptors 3Document27 pagesOrphan Receptors 3Alonso SánchezNo ratings yet
- Corticoides Gen y NogenDocument10 pagesCorticoides Gen y NogenCarlos BerumenNo ratings yet
- 1 s2.0 S0898656821000693 MainDocument4 pages1 s2.0 S0898656821000693 MainLívia MeloNo ratings yet
- Nihms 1873787Document29 pagesNihms 1873787Tibor SzénásiNo ratings yet
- Nature 11896Document10 pagesNature 11896Sameera HameedNo ratings yet
- 5ht2a AgonistsDocument31 pages5ht2a AgoniststhalesNo ratings yet
- Bhattacharya 2004Document5 pagesBhattacharya 2004paul herreraNo ratings yet
- G-Protein-Coupled Receptors and Tyrosine Kinases: Crossroads in Cell Signaling and RegulationDocument7 pagesG-Protein-Coupled Receptors and Tyrosine Kinases: Crossroads in Cell Signaling and RegulationFlorencia FirenzeNo ratings yet
- New Insights Into Modes of GPCR ActivationDocument20 pagesNew Insights Into Modes of GPCR ActivationChasconaNo ratings yet
- 1 IntroductionDocument35 pages1 IntroductionGABRIELA NORMA SOLANO CANCHAYANo ratings yet
- Signal Transduction and Second Messengers: Karen LounsburyDocument10 pagesSignal Transduction and Second Messengers: Karen LounsburyFranco RoaNo ratings yet
- GPCRDocument32 pagesGPCRSergio UribeNo ratings yet
- Review: Molecular Signatures of G-Protein-Coupled ReceptorsDocument10 pagesReview: Molecular Signatures of G-Protein-Coupled ReceptorsJosué VelázquezNo ratings yet
- TMP D19 CDocument124 pagesTMP D19 CFrontiersNo ratings yet
- The Role of Calcitonin Gene-Related Peptide in Peripheral and Central Pain Mechanisms Including MigraineDocument17 pagesThe Role of Calcitonin Gene-Related Peptide in Peripheral and Central Pain Mechanisms Including MigrainegarridotfNo ratings yet
- Reviews: Seven-Transmembrane ReceptorsDocument12 pagesReviews: Seven-Transmembrane Receptorssant_grNo ratings yet
- Señalización Molecular: Julio Hilario VargasDocument43 pagesSeñalización Molecular: Julio Hilario VargasAndrew Emilio Castillo PachecoNo ratings yet
- G Protein Coupled Receptors Research PaperDocument7 pagesG Protein Coupled Receptors Research Paperaflbvmogk100% (1)
- Pnas 202001270Document10 pagesPnas 202001270diego.battiatoNo ratings yet
- Adaptor Protein-2 Interaction With Arrestin Regulates GPCR Recycling and ApoptosisDocument15 pagesAdaptor Protein-2 Interaction With Arrestin Regulates GPCR Recycling and ApoptosisBruna FrancieleNo ratings yet
- New Insights Into Modes of GPCR Activation: ReviewDocument20 pagesNew Insights Into Modes of GPCR Activation: ReviewBenjamin VillagraNo ratings yet
- Depression Rgs8 GenDocument10 pagesDepression Rgs8 GenJOSÉ DAVID SALCEDO CASTRONo ratings yet
- Orphan Receptor GPR158 Serves As A Metabotropic Glycine Receptor mGlyRDocument7 pagesOrphan Receptor GPR158 Serves As A Metabotropic Glycine Receptor mGlyRStress LessNo ratings yet
- Bioorganic & Medicinal Chemistry Letters: Zoran Rankovic, Tarsis F. Brust, Laura M. BohnDocument10 pagesBioorganic & Medicinal Chemistry Letters: Zoran Rankovic, Tarsis F. Brust, Laura M. Bohnkadioglu20No ratings yet
- Jcem 1476Document6 pagesJcem 1476Teodora OnofreiNo ratings yet
- Nair 2017Document14 pagesNair 2017AnkanNo ratings yet
- G-Protein Coupled Receptors (GPCR)Document43 pagesG-Protein Coupled Receptors (GPCR)Avin GupthaNo ratings yet
- GPCR 160211124029Document83 pagesGPCR 160211124029JuhiJahan AmanullahNo ratings yet
- Multiplex GPCR Screen Reveals Reliably Acting Agonists and A Gq-Phospholipase C Coupling Mode of GPR30/GPER1Document57 pagesMultiplex GPCR Screen Reveals Reliably Acting Agonists and A Gq-Phospholipase C Coupling Mode of GPR30/GPER1Benjamin VillagraNo ratings yet
- Corea Del Sur 2019 SODIUMGLUCOSE COTRANSPORTER 2 INHIBOTOR FORDocument1 pageCorea Del Sur 2019 SODIUMGLUCOSE COTRANSPORTER 2 INHIBOTOR FORLaura GarciaNo ratings yet
- t4 Trv120027enratas PDFDocument20 pagest4 Trv120027enratas PDFAlicia Mary PicconeNo ratings yet
- Assignment 2Document2 pagesAssignment 2saras87No ratings yet
- Review Article: The metabolic modulator PGC-1α in cancerDocument14 pagesReview Article: The metabolic modulator PGC-1α in cancerAlIsh BhanDariNo ratings yet
- Pharmacology & Therapeutics: Eugenia V. Gurevich, John J.G. Tesmer, Arcady Mushegian, Vsevolod V. GurevichDocument30 pagesPharmacology & Therapeutics: Eugenia V. Gurevich, John J.G. Tesmer, Arcady Mushegian, Vsevolod V. Gurevichnandhus2227No ratings yet
- 2006-Cancer Research13-Gonadotropin-Releasing Hormone Promotes Ovarian Cancer Cell Invasiveness Through C-Jun NH2-terminal Kinase-Mediated Activation of Matrix Metalloproteinase (MMP) - 2 and MMP-9Document9 pages2006-Cancer Research13-Gonadotropin-Releasing Hormone Promotes Ovarian Cancer Cell Invasiveness Through C-Jun NH2-terminal Kinase-Mediated Activation of Matrix Metalloproteinase (MMP) - 2 and MMP-9shidis1028No ratings yet
- Signal Transduction & G Protein-Coupled Receptors: TopicsDocument33 pagesSignal Transduction & G Protein-Coupled Receptors: TopicsEria MarinaNo ratings yet
- 15G Protein-Coupled ReceptorDocument15 pages15G Protein-Coupled ReceptorZiedTrikiNo ratings yet
- RNA Interference in J774 Macrophages Reveals A Role For Coronin 1 in Mycobacterial Trafficking But Not in Actin-Dependent ProcessesDocument11 pagesRNA Interference in J774 Macrophages Reveals A Role For Coronin 1 in Mycobacterial Trafficking But Not in Actin-Dependent ProcessesAbdulaziz AlharbiNo ratings yet
- 794 FullDocument15 pages794 FullTibor SzénásiNo ratings yet
- Receptors: Types Structure FunctionsDocument44 pagesReceptors: Types Structure Functionsmaham1085No ratings yet
- Navarro 2022Document14 pagesNavarro 2022aldo.guerrero.lopez20No ratings yet
- CompetitionDocument7 pagesCompetitionCarlos LoboNo ratings yet
- Role of Rbm3 Gene in CancerDocument40 pagesRole of Rbm3 Gene in CancerAnkit AgarwalNo ratings yet
- Regulation of Innate Immune Functions by Guanylate-Binding ProteinsDocument9 pagesRegulation of Innate Immune Functions by Guanylate-Binding ProteinsRadwan AlzidanNo ratings yet
- Heteroplasia Osea Progresiva.2015Document12 pagesHeteroplasia Osea Progresiva.2015Panque De vainillaNo ratings yet
- Modi Fication and Functional Inhibition of Regulator of G Protein Signaling 4 (RGS4) by 4 Hydroxy-2-NonenalDocument8 pagesModi Fication and Functional Inhibition of Regulator of G Protein Signaling 4 (RGS4) by 4 Hydroxy-2-NonenalscotNo ratings yet
- Intercellular CommunicationDocument8 pagesIntercellular CommunicationShahran KumarNo ratings yet
- 5163 FullDocument6 pages5163 FullDhirajNo ratings yet
- G Protein Coupled ReceptorsDocument25 pagesG Protein Coupled ReceptorsMirza Shaharyar BaigNo ratings yet
- Sas Hes032 7Document7 pagesSas Hes032 7Jose Melmar Autida AutenticoNo ratings yet
- Bronchitis: Pathology and PathophysiologyDocument4 pagesBronchitis: Pathology and Pathophysiologydwi rina putriNo ratings yet
- Suicide in Teenagers 2001Document6 pagesSuicide in Teenagers 2001Simpson KhooNo ratings yet
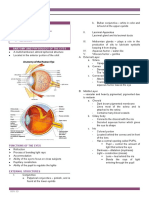
- Eyes Lecture 1Document4 pagesEyes Lecture 1Rue Cheng MaNo ratings yet
- Concept Map PT 1Document1 pageConcept Map PT 1api-657741346No ratings yet
- Radiographic Special Procedures: Kenneth M. Luciano, RRTDocument36 pagesRadiographic Special Procedures: Kenneth M. Luciano, RRTKaye A. JardinicoNo ratings yet
- NURS 6521N Final Exam Question and Answers Feb 2020 (100/100)Document19 pagesNURS 6521N Final Exam Question and Answers Feb 2020 (100/100)Judy Durkin0% (1)
- Obesity and Anticipated Difficult Airway - A Comprehensive Approach With Videolaryngoscopy, Ramp Position, Sevoflurane and Opioid Free AnaesthesiaDocument8 pagesObesity and Anticipated Difficult Airway - A Comprehensive Approach With Videolaryngoscopy, Ramp Position, Sevoflurane and Opioid Free AnaesthesiaIJAR JOURNALNo ratings yet
- p450 Table Oct 11 2009Document2 pagesp450 Table Oct 11 2009Naresh BabuNo ratings yet
- Summary of Product Characteristics: Edronax 4 MGDocument9 pagesSummary of Product Characteristics: Edronax 4 MGddandan_2No ratings yet
- Harvared EditedDocument1 pageHarvared EditedAmirul Assyraf NoorNo ratings yet
- Company ProfileDocument3 pagesCompany ProfileAnmol Sharma Dadhich100% (1)
- Clinical Spectrum of Atherosclerotic Heart DiseaseDocument50 pagesClinical Spectrum of Atherosclerotic Heart Diseaseadamu mohammadNo ratings yet
- Obat Klinik 2022-2023Document96 pagesObat Klinik 2022-2023jeane rahmanNo ratings yet
- Arni PPT 2 PDFDocument19 pagesArni PPT 2 PDFKumarsai Durusoju100% (1)
- Gastrointestinal Hormones Regulating AppetiteDocument23 pagesGastrointestinal Hormones Regulating AppetiteHeba IyadNo ratings yet
- Cef 3Document1 pageCef 3Sabbir AhmedNo ratings yet
- Chapter 23 Lippincott BiochemistryDocument44 pagesChapter 23 Lippincott BiochemistryMeysam SajjadiNo ratings yet
- Anti Diabetic DrugDocument45 pagesAnti Diabetic DrugRahul LokhandeNo ratings yet
- Group 1 Lipid Lowering AgentsDocument70 pagesGroup 1 Lipid Lowering AgentsReylin SaraminesNo ratings yet
- Preoperative EvaluationDocument65 pagesPreoperative Evaluationdr_trialNo ratings yet
- MAR Case 4Document4 pagesMAR Case 4Juviely PremacioNo ratings yet
- Pharma Prelim Assignment 1 - Pharmacodynamics..Document2 pagesPharma Prelim Assignment 1 - Pharmacodynamics..Kimberly Wyne SantosNo ratings yet
- NBDE Remembered 2 New YorkDocument242 pagesNBDE Remembered 2 New YorkR MNo ratings yet
- 731-Article Text-3619-2-10-20210121Document3 pages731-Article Text-3619-2-10-20210121Αβραξας ΓαβριήλNo ratings yet
- Interest of Early Toxicological Sampling During Alpha Chloralose in Toxication, A Case ReportDocument4 pagesInterest of Early Toxicological Sampling During Alpha Chloralose in Toxication, A Case ReportIJAR JOURNALNo ratings yet
- Histamine and Antihistaminic Agents: Pharmaceutical Chemistry 2Document45 pagesHistamine and Antihistaminic Agents: Pharmaceutical Chemistry 2د. أبجًملNo ratings yet
- Alzheimer DiseaseDocument6 pagesAlzheimer DiseaseYousif JawadNo ratings yet
- Hyproliferative Anemias: What Is Anemia?Document15 pagesHyproliferative Anemias: What Is Anemia?yamie sulongNo ratings yet
- Pain 1585622753Document284 pagesPain 1585622753eva pravitasari nefertitiNo ratings yet