
Revision de Celulas Madre Cancerosas Articulo 2
Revision de Celulas Madre Cancerosas Articulo 2
You might also like
- Test Bank For Quick and Easy Medical Terminology 9th Edition by LeonardDocument15 pagesTest Bank For Quick and Easy Medical Terminology 9th Edition by Leonardamyrileyixgnaoqjmt100% (30)
- Basic Mechanisms in Craniofacial GrowthDocument9 pagesBasic Mechanisms in Craniofacial GrowthFabian BarretoNo ratings yet
- Nervous System AlterationsDocument62 pagesNervous System AlterationsAsal SalahNo ratings yet
- Inhibition of Sonic Hedgehog Pathway and Pluripotency Maintaining Factors Regulate Human Pancreatic Cancer Stem Cell CharacteristicsDocument20 pagesInhibition of Sonic Hedgehog Pathway and Pluripotency Maintaining Factors Regulate Human Pancreatic Cancer Stem Cell Characteristicsrakesh srivastavNo ratings yet
- 《文献精读》第4课"肿瘤干细胞" 文献Document20 pages《文献精读》第4课"肿瘤干细胞" 文献zhe zhNo ratings yet
- 1 s2.0 S1044579X22002644 Main 1Document15 pages1 s2.0 S1044579X22002644 Main 1ali69rmrimsNo ratings yet
- Stem Cell Concepts Renew Cancer Research: ASH 50th Anniversary ReviewDocument15 pagesStem Cell Concepts Renew Cancer Research: ASH 50th Anniversary ReviewbloodbenderNo ratings yet
- Pancreatic Cancer Stem CellsDocument9 pagesPancreatic Cancer Stem CellsDaniel RusieNo ratings yet
- Precision Oncology Colorectal CancerDocument23 pagesPrecision Oncology Colorectal CancerNasheen NaidooNo ratings yet
- Pancreas e Cell StaminaliDocument4 pagesPancreas e Cell StaminaliAntonio MichelottiNo ratings yet
- PC3 Is A Cell Line Characteristic of Prostatic Small Cell CarcinomaDocument12 pagesPC3 Is A Cell Line Characteristic of Prostatic Small Cell CarcinomaANGEL SAUL SERRANO ANGULONo ratings yet
- The Identi Fications and Clinical Implications of Cancer Stem Cells in Colorectal CancerDocument10 pagesThe Identi Fications and Clinical Implications of Cancer Stem Cells in Colorectal CancerManuel GuachoNo ratings yet
- Organo TropismDocument12 pagesOrgano TropismKL TongsonNo ratings yet
- Breast Cancer and Cancer Stem Cells: A Mini-Review: Tu Mo Ri, 100: 363-369, 2014Document7 pagesBreast Cancer and Cancer Stem Cells: A Mini-Review: Tu Mo Ri, 100: 363-369, 2014Ava BaranNo ratings yet
- Cancer de Pancreas en RatasDocument8 pagesCancer de Pancreas en RatasAlethia UkweliNo ratings yet
- Cancer y Celulas Madre 2022Document11 pagesCancer y Celulas Madre 2022Jose EdgarNo ratings yet
- NIH Public Access: Author ManuscriptDocument17 pagesNIH Public Access: Author ManuscriptmarcussiNo ratings yet
- Haake Et Al 2017 CancerDocument10 pagesHaake Et Al 2017 Cancerdiego peinadoNo ratings yet
- Understanding and Targeting Cancer Stem Cells Therapeutic Implications and ChallengesDocument9 pagesUnderstanding and Targeting Cancer Stem Cells Therapeutic Implications and ChallengesMariano PerezNo ratings yet
- Geneticsofgastriccancer: Matthew S. Strand,, Albert Craig Lockhart,, Ryan C. FieldsDocument26 pagesGeneticsofgastriccancer: Matthew S. Strand,, Albert Craig Lockhart,, Ryan C. Fieldsjomigp01No ratings yet
- Koshkin 2016Document10 pagesKoshkin 2016chemistpl420No ratings yet
- W de WenisimoDocument18 pagesW de WenisimoBJ CarminatorNo ratings yet
- Cancers 15 00565Document12 pagesCancers 15 00565Sediki ZakariaNo ratings yet
- Polyp Genetics: Coen Laurens Klos, MD Sekhar Dharmarajan, MDDocument7 pagesPolyp Genetics: Coen Laurens Klos, MD Sekhar Dharmarajan, MDMarjuani yusufNo ratings yet
- Primordial Germ Cells As A Potential Shared Cell of Origin For Mucinous Cystic Neoplasms of The Pancreas and Mucinous Ovarian TumorsDocument18 pagesPrimordial Germ Cells As A Potential Shared Cell of Origin For Mucinous Cystic Neoplasms of The Pancreas and Mucinous Ovarian TumorsJagmohanNo ratings yet
- Kurman 2013Document6 pagesKurman 2013adevanshi3399No ratings yet
- CSC Colorectal 6Document9 pagesCSC Colorectal 6Diana LaviniaNo ratings yet
- Cancer Stem Cells ThesisDocument8 pagesCancer Stem Cells Thesisafknekkfs100% (1)
- Cancer-Associated Fibroblasts in Breast Cancer ChallengesDocument34 pagesCancer-Associated Fibroblasts in Breast Cancer ChallengesANA PAULANo ratings yet
- Medsci 06 00031 PDFDocument113 pagesMedsci 06 00031 PDFCarolina VillalobosNo ratings yet
- Creatine Phosphokinase Levels in Oral Cancer Patients: Issn 0974-3618 (Print) 0974-360X (Online)Document4 pagesCreatine Phosphokinase Levels in Oral Cancer Patients: Issn 0974-3618 (Print) 0974-360X (Online)Deepak KumarNo ratings yet
- The Cancer HandbookDocument1,529 pagesThe Cancer HandbookAsimaJulianaSiregarNo ratings yet
- Ni Hms 511528Document19 pagesNi Hms 511528Anonymous PKE8zOXNo ratings yet
- DNA Aptamer Evolved by Cell-SELEX For Recognition ofDocument10 pagesDNA Aptamer Evolved by Cell-SELEX For Recognition ofFernanda LDNo ratings yet
- Review Article: The Recent Advances On Liver Cancer Stem Cells: Biomarkers, Separation, and TherapyDocument10 pagesReview Article: The Recent Advances On Liver Cancer Stem Cells: Biomarkers, Separation, and TherapyNovita ApramadhaNo ratings yet
- Cancer de ColonDocument7 pagesCancer de ColonAlex MoraNo ratings yet
- Jurnal Tumor Metastasis Paru Hepar Dari PancreasDocument15 pagesJurnal Tumor Metastasis Paru Hepar Dari PancreaslucyishakNo ratings yet
- Cancer Stem Cells and Evolving Novel Therapies: A Paradigm ShiftDocument6 pagesCancer Stem Cells and Evolving Novel Therapies: A Paradigm Shifttarry girothNo ratings yet
- Nuevo Forma de Investigar El Cancer PancreaticoDocument8 pagesNuevo Forma de Investigar El Cancer PancreaticovillalobosgarzacarlosfranciscoNo ratings yet
- Metastatic Behavior of Breast Cancer Subtypes: Ournal of Linical NcologyDocument7 pagesMetastatic Behavior of Breast Cancer Subtypes: Ournal of Linical NcologyJafet Gonzalez EstradaNo ratings yet
- Cancer Stem Cells (CSCS) : DR - Amini By: Rouholah Soleimani Amirmohammad SeifDocument20 pagesCancer Stem Cells (CSCS) : DR - Amini By: Rouholah Soleimani Amirmohammad Seifroholla.solimani7820No ratings yet
- 2020 Schcolnik Et AlDocument11 pages2020 Schcolnik Et AlArturo RojoNo ratings yet
- Epistemology of The Origin of Cancer: A New Paradigm: Hypothesis Open AccessDocument15 pagesEpistemology of The Origin of Cancer: A New Paradigm: Hypothesis Open AccessNafiisahRNo ratings yet
- Ovarian Cancer Review - OHNDocument48 pagesOvarian Cancer Review - OHNalejandro.britoaguNo ratings yet
- Cancer and Tumor MicroenvironmentDocument20 pagesCancer and Tumor MicroenvironmentFlorin BranisteNo ratings yet
- Molecular Profiling in Muscle-Invasive Bladder Cancer: More Than The Sum of Its PartsDocument11 pagesMolecular Profiling in Muscle-Invasive Bladder Cancer: More Than The Sum of Its PartsJoel ChávezdrumsNo ratings yet
- A Literature Review of Complementary and Alternative Medicine Use by Colorectal Cancer PatientsDocument4 pagesA Literature Review of Complementary and Alternative Medicine Use by Colorectal Cancer Patientsgw0a869xNo ratings yet
- Liquid Biopsy From Discovery To Clinical ImplementationDocument5 pagesLiquid Biopsy From Discovery To Clinical ImplementationLais BritoNo ratings yet
- Cancer Stem Cells With Increased Metastatic Potential As - 2017 - Seminars in CaDocument7 pagesCancer Stem Cells With Increased Metastatic Potential As - 2017 - Seminars in CaFlorin AchimNo ratings yet
- Prognostic Biomarkers For Survival in Nasopharyngeal Carcinoma - A Systematic Review of The LiteratureDocument14 pagesPrognostic Biomarkers For Survival in Nasopharyngeal Carcinoma - A Systematic Review of The LiteratureWening Dewi HapsariNo ratings yet
- Cancer Stem Cell Hypothesis: Implication For Cancer Prevention and TreatmentDocument16 pagesCancer Stem Cell Hypothesis: Implication For Cancer Prevention and TreatmentlaurentiaNo ratings yet
- Molecular Biomarkers of Bladder Cancer: A Mini-ReviewDocument10 pagesMolecular Biomarkers of Bladder Cancer: A Mini-ReviewaldyNo ratings yet
- Mesothelin Promotes Epithelial-Tomesenchymal Transition and TumorigenicityDocument13 pagesMesothelin Promotes Epithelial-Tomesenchymal Transition and Tumorigenicitypaulo saNo ratings yet
- Deros A 2015Document10 pagesDeros A 2015rakaNo ratings yet
- En - 0120 9957 RCG 33 02 00134 PDFDocument10 pagesEn - 0120 9957 RCG 33 02 00134 PDFbernardo04061993No ratings yet
- Cancer Cells Research PaperDocument4 pagesCancer Cells Research Paperfusolemebeg2100% (1)
- Prognostic Significance of PINCH Signalling in Human Pancreatic Ductal AdenocarcinomaDocument7 pagesPrognostic Significance of PINCH Signalling in Human Pancreatic Ductal AdenocarcinomaLuis FuentesNo ratings yet
- Langer 2010Document10 pagesLanger 2010Nelson William UsnayoNo ratings yet
- NEJMhereditarycolorectalcancer PDFDocument14 pagesNEJMhereditarycolorectalcancer PDF송란다No ratings yet
- PG Art 34580-10Document15 pagesPG Art 34580-10Horia MineaNo ratings yet
- 10 Rosenberg Vitamin CDocument78 pages10 Rosenberg Vitamin CЩербакова ЛенаNo ratings yet
- Carcinoma of PancreasDocument15 pagesCarcinoma of Pancreasعمار عارفNo ratings yet
- Abscesses Lecture - Physiotherapists. 2012Document24 pagesAbscesses Lecture - Physiotherapists. 2012divinaNo ratings yet
- The Meninges - Dura - Arachnoid - Pia - TeachMeAnatomyDocument4 pagesThe Meninges - Dura - Arachnoid - Pia - TeachMeAnatomyraihaana turawaNo ratings yet
- Hypoxic-Ischemic Encephalopathy in Adult: January 2010Document5 pagesHypoxic-Ischemic Encephalopathy in Adult: January 2010Pasien ICU RSUINo ratings yet
- 12th Bio - Zoology Unit Wise 1 Mark Study MaterialDocument29 pages12th Bio - Zoology Unit Wise 1 Mark Study Materialfredrick100% (1)
- Stroke: Pembimbing: Dr. Handojo Pudjowidyanto, SpsDocument88 pagesStroke: Pembimbing: Dr. Handojo Pudjowidyanto, SpsWahyu FajarNo ratings yet
- Atlas of Spleen Pathology and Histhology CompletDocument173 pagesAtlas of Spleen Pathology and Histhology CompletAncamed NycoletaNo ratings yet
- Depolarizing Muscle RelaxantDocument40 pagesDepolarizing Muscle RelaxantMohammad HayajnehNo ratings yet
- Programmed Cell Death in AgingDocument25 pagesProgrammed Cell Death in AgingGustavo Sá MottaNo ratings yet
- Full Download PDF of Revision Lumbar Spine Surgery 1st Edition Robert F. Heary - Ebook PDF All ChapterDocument69 pagesFull Download PDF of Revision Lumbar Spine Surgery 1st Edition Robert F. Heary - Ebook PDF All Chaptervockoairike100% (7)
- Partial Hydatidiform Mole: Notre Dame of Marbel UniversityDocument127 pagesPartial Hydatidiform Mole: Notre Dame of Marbel UniversityInsatiable CleeNo ratings yet
- Autopsy Manual: NAVM ED P-5065 AFM 160-19Document130 pagesAutopsy Manual: NAVM ED P-5065 AFM 160-19Roger ChristensenNo ratings yet
- 111 Peripheral NeuropathyDocument17 pages111 Peripheral NeuropathyjassbhanguNo ratings yet
- Detecting and Recognising Lung Cancer: Using Convolutional Neural NetworksDocument25 pagesDetecting and Recognising Lung Cancer: Using Convolutional Neural NetworksRAJU MAURYANo ratings yet
- Module 2 - Developing The Whole PersonDocument91 pagesModule 2 - Developing The Whole PersonArchi Archi100% (2)
- The Neuro Ophthalmology Survival Guide Second Edition Edition Burdon Ebook Full ChapterDocument51 pagesThe Neuro Ophthalmology Survival Guide Second Edition Edition Burdon Ebook Full Chapterjuan.small935100% (20)
- Cardiovascular Physiology 0 Anesthetic ImportanceDocument100 pagesCardiovascular Physiology 0 Anesthetic ImportanceSurya SuryaNo ratings yet
- MCQ Final 1800Document19 pagesMCQ Final 1800JohnSonNo ratings yet
- 2023 The Sanford Guide - PDFDocument40 pages2023 The Sanford Guide - PDFMinodora FaurNo ratings yet
- Neurological Examination Made Easy 6E 6Th Edition Geraint Fuller Ma MD FRCP Full ChapterDocument53 pagesNeurological Examination Made Easy 6E 6Th Edition Geraint Fuller Ma MD FRCP Full Chapterlinda.elgin584100% (12)
- Urinary Renal System - 4th EdDocument31 pagesUrinary Renal System - 4th EdpadialmichelleNo ratings yet
- Ramji 2015Document13 pagesRamji 2015AlexandraNo ratings yet
- SALIVARY GLAND IMAGING-Seminar ConversionDocument114 pagesSALIVARY GLAND IMAGING-Seminar ConversionVanshika JainNo ratings yet
- Jurnal 11Document7 pagesJurnal 11Zella ZakyaNo ratings yet
- Ervous Ystem: Structural ClassificationDocument95 pagesErvous Ystem: Structural ClassificationJobelle AcenaNo ratings yet
- Nervous SystemDocument79 pagesNervous SystemAffan ElahiNo ratings yet
- Oral Pathology (Review Center)Document12 pagesOral Pathology (Review Center)yellow rangerNo ratings yet































































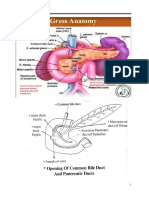

























Download as pdf or txt
You might also like
- Test Bank For Quick and Easy Medical Terminology 9th Edition by LeonardDocument15 pagesTest Bank For Quick and Easy Medical Terminology 9th Edition by Leonardamyrileyixgnaoqjmt100% (30)
- Basic Mechanisms in Craniofacial GrowthDocument9 pagesBasic Mechanisms in Craniofacial GrowthFabian BarretoNo ratings yet
- Nervous System AlterationsDocument62 pagesNervous System AlterationsAsal SalahNo ratings yet
- Inhibition of Sonic Hedgehog Pathway and Pluripotency Maintaining Factors Regulate Human Pancreatic Cancer Stem Cell CharacteristicsDocument20 pagesInhibition of Sonic Hedgehog Pathway and Pluripotency Maintaining Factors Regulate Human Pancreatic Cancer Stem Cell Characteristicsrakesh srivastavNo ratings yet
- 《文献精读》第4课"肿瘤干细胞" 文献Document20 pages《文献精读》第4课"肿瘤干细胞" 文献zhe zhNo ratings yet
- 1 s2.0 S1044579X22002644 Main 1Document15 pages1 s2.0 S1044579X22002644 Main 1ali69rmrimsNo ratings yet
- Stem Cell Concepts Renew Cancer Research: ASH 50th Anniversary ReviewDocument15 pagesStem Cell Concepts Renew Cancer Research: ASH 50th Anniversary ReviewbloodbenderNo ratings yet
- Pancreatic Cancer Stem CellsDocument9 pagesPancreatic Cancer Stem CellsDaniel RusieNo ratings yet
- Precision Oncology Colorectal CancerDocument23 pagesPrecision Oncology Colorectal CancerNasheen NaidooNo ratings yet
- Pancreas e Cell StaminaliDocument4 pagesPancreas e Cell StaminaliAntonio MichelottiNo ratings yet
- PC3 Is A Cell Line Characteristic of Prostatic Small Cell CarcinomaDocument12 pagesPC3 Is A Cell Line Characteristic of Prostatic Small Cell CarcinomaANGEL SAUL SERRANO ANGULONo ratings yet
- The Identi Fications and Clinical Implications of Cancer Stem Cells in Colorectal CancerDocument10 pagesThe Identi Fications and Clinical Implications of Cancer Stem Cells in Colorectal CancerManuel GuachoNo ratings yet
- Organo TropismDocument12 pagesOrgano TropismKL TongsonNo ratings yet
- Breast Cancer and Cancer Stem Cells: A Mini-Review: Tu Mo Ri, 100: 363-369, 2014Document7 pagesBreast Cancer and Cancer Stem Cells: A Mini-Review: Tu Mo Ri, 100: 363-369, 2014Ava BaranNo ratings yet
- Cancer de Pancreas en RatasDocument8 pagesCancer de Pancreas en RatasAlethia UkweliNo ratings yet
- Cancer y Celulas Madre 2022Document11 pagesCancer y Celulas Madre 2022Jose EdgarNo ratings yet
- NIH Public Access: Author ManuscriptDocument17 pagesNIH Public Access: Author ManuscriptmarcussiNo ratings yet
- Haake Et Al 2017 CancerDocument10 pagesHaake Et Al 2017 Cancerdiego peinadoNo ratings yet
- Understanding and Targeting Cancer Stem Cells Therapeutic Implications and ChallengesDocument9 pagesUnderstanding and Targeting Cancer Stem Cells Therapeutic Implications and ChallengesMariano PerezNo ratings yet
- Geneticsofgastriccancer: Matthew S. Strand,, Albert Craig Lockhart,, Ryan C. FieldsDocument26 pagesGeneticsofgastriccancer: Matthew S. Strand,, Albert Craig Lockhart,, Ryan C. Fieldsjomigp01No ratings yet
- Koshkin 2016Document10 pagesKoshkin 2016chemistpl420No ratings yet
- W de WenisimoDocument18 pagesW de WenisimoBJ CarminatorNo ratings yet
- Cancers 15 00565Document12 pagesCancers 15 00565Sediki ZakariaNo ratings yet
- Polyp Genetics: Coen Laurens Klos, MD Sekhar Dharmarajan, MDDocument7 pagesPolyp Genetics: Coen Laurens Klos, MD Sekhar Dharmarajan, MDMarjuani yusufNo ratings yet
- Primordial Germ Cells As A Potential Shared Cell of Origin For Mucinous Cystic Neoplasms of The Pancreas and Mucinous Ovarian TumorsDocument18 pagesPrimordial Germ Cells As A Potential Shared Cell of Origin For Mucinous Cystic Neoplasms of The Pancreas and Mucinous Ovarian TumorsJagmohanNo ratings yet
- Kurman 2013Document6 pagesKurman 2013adevanshi3399No ratings yet
- CSC Colorectal 6Document9 pagesCSC Colorectal 6Diana LaviniaNo ratings yet
- Cancer Stem Cells ThesisDocument8 pagesCancer Stem Cells Thesisafknekkfs100% (1)
- Cancer-Associated Fibroblasts in Breast Cancer ChallengesDocument34 pagesCancer-Associated Fibroblasts in Breast Cancer ChallengesANA PAULANo ratings yet
- Medsci 06 00031 PDFDocument113 pagesMedsci 06 00031 PDFCarolina VillalobosNo ratings yet
- Creatine Phosphokinase Levels in Oral Cancer Patients: Issn 0974-3618 (Print) 0974-360X (Online)Document4 pagesCreatine Phosphokinase Levels in Oral Cancer Patients: Issn 0974-3618 (Print) 0974-360X (Online)Deepak KumarNo ratings yet
- The Cancer HandbookDocument1,529 pagesThe Cancer HandbookAsimaJulianaSiregarNo ratings yet
- Ni Hms 511528Document19 pagesNi Hms 511528Anonymous PKE8zOXNo ratings yet
- DNA Aptamer Evolved by Cell-SELEX For Recognition ofDocument10 pagesDNA Aptamer Evolved by Cell-SELEX For Recognition ofFernanda LDNo ratings yet
- Review Article: The Recent Advances On Liver Cancer Stem Cells: Biomarkers, Separation, and TherapyDocument10 pagesReview Article: The Recent Advances On Liver Cancer Stem Cells: Biomarkers, Separation, and TherapyNovita ApramadhaNo ratings yet
- Cancer de ColonDocument7 pagesCancer de ColonAlex MoraNo ratings yet
- Jurnal Tumor Metastasis Paru Hepar Dari PancreasDocument15 pagesJurnal Tumor Metastasis Paru Hepar Dari PancreaslucyishakNo ratings yet
- Cancer Stem Cells and Evolving Novel Therapies: A Paradigm ShiftDocument6 pagesCancer Stem Cells and Evolving Novel Therapies: A Paradigm Shifttarry girothNo ratings yet
- Nuevo Forma de Investigar El Cancer PancreaticoDocument8 pagesNuevo Forma de Investigar El Cancer PancreaticovillalobosgarzacarlosfranciscoNo ratings yet
- Metastatic Behavior of Breast Cancer Subtypes: Ournal of Linical NcologyDocument7 pagesMetastatic Behavior of Breast Cancer Subtypes: Ournal of Linical NcologyJafet Gonzalez EstradaNo ratings yet
- Cancer Stem Cells (CSCS) : DR - Amini By: Rouholah Soleimani Amirmohammad SeifDocument20 pagesCancer Stem Cells (CSCS) : DR - Amini By: Rouholah Soleimani Amirmohammad Seifroholla.solimani7820No ratings yet
- 2020 Schcolnik Et AlDocument11 pages2020 Schcolnik Et AlArturo RojoNo ratings yet
- Epistemology of The Origin of Cancer: A New Paradigm: Hypothesis Open AccessDocument15 pagesEpistemology of The Origin of Cancer: A New Paradigm: Hypothesis Open AccessNafiisahRNo ratings yet
- Ovarian Cancer Review - OHNDocument48 pagesOvarian Cancer Review - OHNalejandro.britoaguNo ratings yet
- Cancer and Tumor MicroenvironmentDocument20 pagesCancer and Tumor MicroenvironmentFlorin BranisteNo ratings yet
- Molecular Profiling in Muscle-Invasive Bladder Cancer: More Than The Sum of Its PartsDocument11 pagesMolecular Profiling in Muscle-Invasive Bladder Cancer: More Than The Sum of Its PartsJoel ChávezdrumsNo ratings yet
- A Literature Review of Complementary and Alternative Medicine Use by Colorectal Cancer PatientsDocument4 pagesA Literature Review of Complementary and Alternative Medicine Use by Colorectal Cancer Patientsgw0a869xNo ratings yet
- Liquid Biopsy From Discovery To Clinical ImplementationDocument5 pagesLiquid Biopsy From Discovery To Clinical ImplementationLais BritoNo ratings yet
- Cancer Stem Cells With Increased Metastatic Potential As - 2017 - Seminars in CaDocument7 pagesCancer Stem Cells With Increased Metastatic Potential As - 2017 - Seminars in CaFlorin AchimNo ratings yet
- Prognostic Biomarkers For Survival in Nasopharyngeal Carcinoma - A Systematic Review of The LiteratureDocument14 pagesPrognostic Biomarkers For Survival in Nasopharyngeal Carcinoma - A Systematic Review of The LiteratureWening Dewi HapsariNo ratings yet
- Cancer Stem Cell Hypothesis: Implication For Cancer Prevention and TreatmentDocument16 pagesCancer Stem Cell Hypothesis: Implication For Cancer Prevention and TreatmentlaurentiaNo ratings yet
- Molecular Biomarkers of Bladder Cancer: A Mini-ReviewDocument10 pagesMolecular Biomarkers of Bladder Cancer: A Mini-ReviewaldyNo ratings yet
- Mesothelin Promotes Epithelial-Tomesenchymal Transition and TumorigenicityDocument13 pagesMesothelin Promotes Epithelial-Tomesenchymal Transition and Tumorigenicitypaulo saNo ratings yet
- Deros A 2015Document10 pagesDeros A 2015rakaNo ratings yet
- En - 0120 9957 RCG 33 02 00134 PDFDocument10 pagesEn - 0120 9957 RCG 33 02 00134 PDFbernardo04061993No ratings yet
- Cancer Cells Research PaperDocument4 pagesCancer Cells Research Paperfusolemebeg2100% (1)
- Prognostic Significance of PINCH Signalling in Human Pancreatic Ductal AdenocarcinomaDocument7 pagesPrognostic Significance of PINCH Signalling in Human Pancreatic Ductal AdenocarcinomaLuis FuentesNo ratings yet
- Langer 2010Document10 pagesLanger 2010Nelson William UsnayoNo ratings yet
- NEJMhereditarycolorectalcancer PDFDocument14 pagesNEJMhereditarycolorectalcancer PDF송란다No ratings yet
- PG Art 34580-10Document15 pagesPG Art 34580-10Horia MineaNo ratings yet
- 10 Rosenberg Vitamin CDocument78 pages10 Rosenberg Vitamin CЩербакова ЛенаNo ratings yet
- Carcinoma of PancreasDocument15 pagesCarcinoma of Pancreasعمار عارفNo ratings yet
- Abscesses Lecture - Physiotherapists. 2012Document24 pagesAbscesses Lecture - Physiotherapists. 2012divinaNo ratings yet
- The Meninges - Dura - Arachnoid - Pia - TeachMeAnatomyDocument4 pagesThe Meninges - Dura - Arachnoid - Pia - TeachMeAnatomyraihaana turawaNo ratings yet
- Hypoxic-Ischemic Encephalopathy in Adult: January 2010Document5 pagesHypoxic-Ischemic Encephalopathy in Adult: January 2010Pasien ICU RSUINo ratings yet
- 12th Bio - Zoology Unit Wise 1 Mark Study MaterialDocument29 pages12th Bio - Zoology Unit Wise 1 Mark Study Materialfredrick100% (1)
- Stroke: Pembimbing: Dr. Handojo Pudjowidyanto, SpsDocument88 pagesStroke: Pembimbing: Dr. Handojo Pudjowidyanto, SpsWahyu FajarNo ratings yet
- Atlas of Spleen Pathology and Histhology CompletDocument173 pagesAtlas of Spleen Pathology and Histhology CompletAncamed NycoletaNo ratings yet
- Depolarizing Muscle RelaxantDocument40 pagesDepolarizing Muscle RelaxantMohammad HayajnehNo ratings yet
- Programmed Cell Death in AgingDocument25 pagesProgrammed Cell Death in AgingGustavo Sá MottaNo ratings yet
- Full Download PDF of Revision Lumbar Spine Surgery 1st Edition Robert F. Heary - Ebook PDF All ChapterDocument69 pagesFull Download PDF of Revision Lumbar Spine Surgery 1st Edition Robert F. Heary - Ebook PDF All Chaptervockoairike100% (7)
- Partial Hydatidiform Mole: Notre Dame of Marbel UniversityDocument127 pagesPartial Hydatidiform Mole: Notre Dame of Marbel UniversityInsatiable CleeNo ratings yet
- Autopsy Manual: NAVM ED P-5065 AFM 160-19Document130 pagesAutopsy Manual: NAVM ED P-5065 AFM 160-19Roger ChristensenNo ratings yet
- 111 Peripheral NeuropathyDocument17 pages111 Peripheral NeuropathyjassbhanguNo ratings yet
- Detecting and Recognising Lung Cancer: Using Convolutional Neural NetworksDocument25 pagesDetecting and Recognising Lung Cancer: Using Convolutional Neural NetworksRAJU MAURYANo ratings yet
- Module 2 - Developing The Whole PersonDocument91 pagesModule 2 - Developing The Whole PersonArchi Archi100% (2)
- The Neuro Ophthalmology Survival Guide Second Edition Edition Burdon Ebook Full ChapterDocument51 pagesThe Neuro Ophthalmology Survival Guide Second Edition Edition Burdon Ebook Full Chapterjuan.small935100% (20)
- Cardiovascular Physiology 0 Anesthetic ImportanceDocument100 pagesCardiovascular Physiology 0 Anesthetic ImportanceSurya SuryaNo ratings yet
- MCQ Final 1800Document19 pagesMCQ Final 1800JohnSonNo ratings yet
- 2023 The Sanford Guide - PDFDocument40 pages2023 The Sanford Guide - PDFMinodora FaurNo ratings yet
- Neurological Examination Made Easy 6E 6Th Edition Geraint Fuller Ma MD FRCP Full ChapterDocument53 pagesNeurological Examination Made Easy 6E 6Th Edition Geraint Fuller Ma MD FRCP Full Chapterlinda.elgin584100% (12)
- Urinary Renal System - 4th EdDocument31 pagesUrinary Renal System - 4th EdpadialmichelleNo ratings yet
- Ramji 2015Document13 pagesRamji 2015AlexandraNo ratings yet
- SALIVARY GLAND IMAGING-Seminar ConversionDocument114 pagesSALIVARY GLAND IMAGING-Seminar ConversionVanshika JainNo ratings yet
- Jurnal 11Document7 pagesJurnal 11Zella ZakyaNo ratings yet
- Ervous Ystem: Structural ClassificationDocument95 pagesErvous Ystem: Structural ClassificationJobelle AcenaNo ratings yet
- Nervous SystemDocument79 pagesNervous SystemAffan ElahiNo ratings yet
- Oral Pathology (Review Center)Document12 pagesOral Pathology (Review Center)yellow rangerNo ratings yet