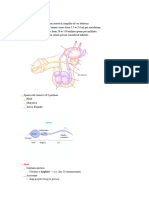
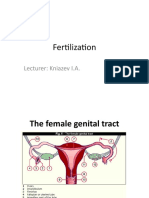
Download as pdf or txt
You might also like
- An Examination of Children S Senses The Damaging Effects of Media and The Therapeutic Possibilities of Puppetry PDFDocument121 pagesAn Examination of Children S Senses The Damaging Effects of Media and The Therapeutic Possibilities of Puppetry PDFsiniordottoreNo ratings yet
- AlakeshBarman Zoology PG 2ND FERTILIZATIONDocument28 pagesAlakeshBarman Zoology PG 2ND FERTILIZATIONTrueAlpha 16No ratings yet
- HIMB Sea Urchin Fertilization LabDocument15 pagesHIMB Sea Urchin Fertilization LabAmy HollingsworthNo ratings yet
- FertilizationDocument10 pagesFertilizationdeepakshi rajputNo ratings yet
- Alakeshbarman Zoology PG 2nd FertilizationDocument28 pagesAlakeshbarman Zoology PG 2nd Fertilizationouafia liiamaniNo ratings yet
- 1st WeekDocument25 pages1st WeekNamra NoorNo ratings yet
- 100Document12 pages100Rajat K. SNo ratings yet
- Acrosome ReactionDocument4 pagesAcrosome ReactionKuldip DwivediNo ratings yet
- Fertilization: Beginning A New Organism: Lee Kui SoonDocument31 pagesFertilization: Beginning A New Organism: Lee Kui SoonSayapPutihNo ratings yet
- HumanDocument18 pagesHumanDavidson VishalNo ratings yet
- Gamete Fusion and The Prevention of PolyspermyDocument9 pagesGamete Fusion and The Prevention of Polyspermyme and allNo ratings yet
- Biol 216 2020 Topic 2 Development - Powerpoint 2.0 Fertilization - TaggedDocument24 pagesBiol 216 2020 Topic 2 Development - Powerpoint 2.0 Fertilization - TaggedMichael HamoudiNo ratings yet
- Embryology - Fertilization: Paper 1 - Part BDocument5 pagesEmbryology - Fertilization: Paper 1 - Part BViswa GiriNo ratings yet
- Prof. Dr. Çiğdem ElmasDocument49 pagesProf. Dr. Çiğdem ElmasMuhammet Fatih CantepeNo ratings yet
- Chapter 1 Part 3 FertilizationDocument16 pagesChapter 1 Part 3 FertilizationSyeda Fizza BatoolNo ratings yet
- L-3human Reproduction NeetDocument12 pagesL-3human Reproduction NeetShorya GuptaNo ratings yet
- Human FertilizationDocument3 pagesHuman Fertilizationtamalbio7No ratings yet
- 4 FertilizationDocument59 pages4 FertilizationQaiser InayatNo ratings yet
- FertilizationDocument8 pagesFertilizationAmy WuNo ratings yet
- Reproduction PPT 6 PHYSIOLOGY OF PREGNANCYDocument98 pagesReproduction PPT 6 PHYSIOLOGY OF PREGNANCYlisanames.23No ratings yet
- Class 12 BDocument11 pagesClass 12 BHarsha AntonyNo ratings yet
- Fertilization ClassDocument41 pagesFertilization ClassredoxtrpNo ratings yet
- Human Reproduction-IiDocument11 pagesHuman Reproduction-IiSuryansh SharmaNo ratings yet
- Lec 3 Gametogenesis and FertilizationDocument43 pagesLec 3 Gametogenesis and FertilizationDacks WangNo ratings yet
- Gametes and FertilizationDocument3 pagesGametes and FertilizationCrystal Ann B. LaresmaNo ratings yet
- A BeginningDocument12 pagesA BeginningAmey JatharNo ratings yet
- Phases of Embryonic DevelopmentDocument9 pagesPhases of Embryonic DevelopmentIshu ChoudharyNo ratings yet
- 02 FertilizationDocument6 pages02 Fertilizationhiba jasimNo ratings yet
- Fertilization Overview: Sperm PenetrationDocument13 pagesFertilization Overview: Sperm PenetrationAmira FahruddinNo ratings yet
- Chapter FertilizationDocument6 pagesChapter FertilizationAbd EloihedNo ratings yet
- 8673-2 Doc2Document13 pages8673-2 Doc2Imaan Khan ImaanNo ratings yet
- Short Report: Student: Tran Minh Tung Department of Animal ScienceDocument1 pageShort Report: Student: Tran Minh Tung Department of Animal ScienceTungTranNo ratings yet
- Fertilizacion RevisionDocument8 pagesFertilizacion RevisionAsistente IRP RESTREPONo ratings yet
- FertilisasiDocument10 pagesFertilisasimaya fathurrahmiNo ratings yet
- Module 5Document10 pagesModule 5Phan MhiveNo ratings yet
- Reproduction and Embryonic DevelopmentDocument37 pagesReproduction and Embryonic DevelopmentFXJNo ratings yet
- Human Development Days 1-4Document43 pagesHuman Development Days 1-4Kriztine Mae GonzalesNo ratings yet
- Cellular Mechanism of Sexual ReproductionDocument20 pagesCellular Mechanism of Sexual ReproductionJessa BelleNo ratings yet
- Conception 2021Document15 pagesConception 2021Dhinesh ManoharanNo ratings yet
- Legends of The Sperm and EggDocument8 pagesLegends of The Sperm and EggALNo ratings yet
- Animal - Development (Kel 1-5)Document83 pagesAnimal - Development (Kel 1-5)MelatiNo ratings yet
- Lesson4 FertilizationDocument33 pagesLesson4 Fertilizationy041087No ratings yet
- Makalah Fisiologi HewanDocument11 pagesMakalah Fisiologi HewanMethaNurIndahNo ratings yet
- FertilizationDocument35 pagesFertilizationAlpha GamingNo ratings yet
- Principles of Development: Developmental BiologyDocument86 pagesPrinciples of Development: Developmental BiologyPaolo OcampoNo ratings yet
- Reproduction Part 2Document12 pagesReproduction Part 2Yuh moddaNo ratings yet
- Topic: Mechanisms of Fertilization:: Presented byDocument16 pagesTopic: Mechanisms of Fertilization:: Presented byMajani RajbonshiNo ratings yet
- Animal Reproduction Notes-16Document21 pagesAnimal Reproduction Notes-16manshibhaskar09No ratings yet
- Gametes, Gametogenesis Usually Starts With GametogoniaDocument9 pagesGametes, Gametogenesis Usually Starts With GametogoniaDAGUIBIG, Patricia S.No ratings yet
- PolyspermyDocument2 pagesPolyspermyamnakhanak4748No ratings yet
- Topic: Fertilization: Print Self Learning MaterialDocument9 pagesTopic: Fertilization: Print Self Learning MaterialNihalNo ratings yet
- GA&E 13 - Fertilization & ImplantationDocument49 pagesGA&E 13 - Fertilization & ImplantationSu ZikaiNo ratings yet
- Human Fertilization - Wikipedia, The Free EncyclopediaDocument6 pagesHuman Fertilization - Wikipedia, The Free EncyclopediaNadiya AfifahNo ratings yet
- Fertilization: Recommended Reading: SummaryDocument10 pagesFertilization: Recommended Reading: SummaryKnjigeNo ratings yet
- Semen AnalysisDocument35 pagesSemen AnalysisSalman KhanNo ratings yet
- FertilizationDocument17 pagesFertilizationJahanvi PatelNo ratings yet
- Useful Notes On Fetoplacental Unit and Its Physiological SignificanceDocument3 pagesUseful Notes On Fetoplacental Unit and Its Physiological Significancedr sadia nusrat alamgirNo ratings yet
- Introduction To FertilizationDocument4 pagesIntroduction To FertilizationSupermarc OlaguirNo ratings yet
- IsaacDocument20 pagesIsaacSerein DaleonNo ratings yet
- Developmental Biology: LS 311/ 411 External FertilizationDocument48 pagesDevelopmental Biology: LS 311/ 411 External FertilizationSankalpa ChakrabortyNo ratings yet
- II.A. Geology of Bangsamoro Autonomous Region in Muslim Mindanao (BARMM) Former ArmmDocument3 pagesII.A. Geology of Bangsamoro Autonomous Region in Muslim Mindanao (BARMM) Former ArmmMatthew Julius TatelNo ratings yet
- WS01 Slopes Samantha SarahDocument7 pagesWS01 Slopes Samantha SarahSAMANTHA SARAH PURBANo ratings yet
- Hero Project Kedar BBADocument49 pagesHero Project Kedar BBAVinod HeggannaNo ratings yet
- LTQ Orbitrap Discovery HardwareDocument118 pagesLTQ Orbitrap Discovery Hardwarepribadi socojatiNo ratings yet
- Enprep - 220EC Steel Cu Brass MSDS.Document5 pagesEnprep - 220EC Steel Cu Brass MSDS.LựuLiềuLìNo ratings yet
- Personality DevelopmentDocument7 pagesPersonality DevelopmentArlyn Jane GregorioNo ratings yet
- Perfect Polymers - Product-Introduction - SUNPAL Multiwall Cell) Polycarbonate Standing-Seam Architectural SystemDocument28 pagesPerfect Polymers - Product-Introduction - SUNPAL Multiwall Cell) Polycarbonate Standing-Seam Architectural SystemGirish DhawanNo ratings yet
- PG Prospectus 2024-25 - 23.04.2024Document38 pagesPG Prospectus 2024-25 - 23.04.2024alihossain armanNo ratings yet
- Unit-6 Flow Through PipesDocument54 pagesUnit-6 Flow Through PipesS PrathebaNo ratings yet
- ATR100 Owners Manual PDFDocument20 pagesATR100 Owners Manual PDFHernan Alberto Orozco CastañoNo ratings yet
- Gea-May1-2014flatplate-List-Price-Catalog Plate Heat ExchangerDocument16 pagesGea-May1-2014flatplate-List-Price-Catalog Plate Heat ExchangerpogisimpatikoNo ratings yet
- Manual Reloj DGT 2010 PDFDocument70 pagesManual Reloj DGT 2010 PDFGUILLERMO JLTNo ratings yet
- Calibrationcahrt-Tank KL-10,15,20,50,70Document14 pagesCalibrationcahrt-Tank KL-10,15,20,50,70Uttrsh KrNo ratings yet
- Lab Session 2Document7 pagesLab Session 2usamaNo ratings yet
- PT WBK 777e Kdp00831 Wht019 Smu 5217 Seal Yoke LeakDocument1 pagePT WBK 777e Kdp00831 Wht019 Smu 5217 Seal Yoke LeakNasirKodratNo ratings yet
- Code PICDocument6 pagesCode PICsongbao527No ratings yet
- Meq 01 2015 0012Document17 pagesMeq 01 2015 0012Anonymous Ja3jvaXNo ratings yet
- Vervece Brochure CompactDocument2 pagesVervece Brochure Compacthilmy yusuf maulanaNo ratings yet
- Aegis Basics of MES WhitepaperDocument19 pagesAegis Basics of MES Whitepaperg_325899365100% (1)
- 17 C ENGG NR-1 FATEHGARH-III EQP 400kV ISO STRDocument1 page17 C ENGG NR-1 FATEHGARH-III EQP 400kV ISO STRsarat mishraNo ratings yet
- GENERALISATIONDocument17 pagesGENERALISATIONgreyproperties88No ratings yet
- Reading Comprehension Tests - 83653Document2 pagesReading Comprehension Tests - 83653Henry Rojas VasquezNo ratings yet
- Toxicological Profiles AmmoniaDocument248 pagesToxicological Profiles AmmoniaHugh MantaNo ratings yet
- ACOS730 Flyer 201707 ENDocument5 pagesACOS730 Flyer 201707 ENSentaNo ratings yet
- Cathodic Protection CatalogDocument18 pagesCathodic Protection CatalogfarukNo ratings yet
- Case Study Orientation SlidesDocument88 pagesCase Study Orientation SlidesJoseph Emmanuel CandaNo ratings yet
- Ielts Reading Exercise TypesDocument44 pagesIelts Reading Exercise TypesTú VõNo ratings yet
- Belt Conveyor Idler: Read These Instructions Carefully Before Starting InstallationDocument8 pagesBelt Conveyor Idler: Read These Instructions Carefully Before Starting InstallationKenji LacambraNo ratings yet
- Admission Notification: 3 Floor, Composite Arts Building, Golapbag Golapbag, Burdwan: - 713 104 West Bengal, IndiaDocument3 pagesAdmission Notification: 3 Floor, Composite Arts Building, Golapbag Golapbag, Burdwan: - 713 104 West Bengal, IndiaHemnath Patra দর্শনNo ratings yet