
Purwwasena 2009
Purwwasena 2009
You might also like
- Housing For Swine ProductionDocument87 pagesHousing For Swine ProductionMaria Leonora Theresa100% (3)
- Robert D. Blevins - Flow-Induced Vibration (2001, Krieger Pub Co) PDFDocument254 pagesRobert D. Blevins - Flow-Induced Vibration (2001, Krieger Pub Co) PDFDang Dinh DongNo ratings yet
- Galaxy at War - Star Wars SagaDocument225 pagesGalaxy at War - Star Wars SagaSebastián Rigoni100% (7)
- How To Sell Without Selling - Step - by - Step Marketing Formula To Attract Ready - To - Buy Clients Create Passive Income and Make More Money While Making A DifferenceDocument287 pagesHow To Sell Without Selling - Step - by - Step Marketing Formula To Attract Ready - To - Buy Clients Create Passive Income and Make More Money While Making A Differencedebanikg100% (7)
- Improving Internal Control A Practical Guide For Microfinance InstitutionsDocument83 pagesImproving Internal Control A Practical Guide For Microfinance InstitutionsIon Cara50% (2)
- 2016 Al-Bahry Article - PotentialInHeavyOilBiodegradatDocument13 pages2016 Al-Bahry Article - PotentialInHeavyOilBiodegradatFaqiangNo ratings yet
- Preparation, Characterization and Application of Zeolite-Based Catalyst For Production of Biodiesel From Waste Cooking OilDocument5 pagesPreparation, Characterization and Application of Zeolite-Based Catalyst For Production of Biodiesel From Waste Cooking OilFakhruddin HarahapNo ratings yet
- Determination of Physico-Chemical Properties of Castor Biodiesel A PotentialDocument7 pagesDetermination of Physico-Chemical Properties of Castor Biodiesel A PotentialIAEME PublicationNo ratings yet
- Gassara Et AlDocument6 pagesGassara Et Almushtaq521No ratings yet
- Physico-Chemical Characterization of Algal Oil - Kumar-2011 PDFDocument5 pagesPhysico-Chemical Characterization of Algal Oil - Kumar-2011 PDFrinkanNo ratings yet
- SPE 123303 Bio-Genetic Engineering, Future of Multi Microbial Culture EOR - A Detailed ReportDocument5 pagesSPE 123303 Bio-Genetic Engineering, Future of Multi Microbial Culture EOR - A Detailed ReportSajad FalahNo ratings yet
- Aceite de FrituraDocument8 pagesAceite de FrituraNestor Andres Urbina SuarezNo ratings yet
- Jurnal OkeDocument6 pagesJurnal OkeIlm NafsiNo ratings yet
- 2014 Raed Bacterial Biodegradation of Crude Oil Using LocalDocument8 pages2014 Raed Bacterial Biodegradation of Crude Oil Using LocalFaqiangNo ratings yet
- Production of Sunflower Oil Methyl Esters by Optimized Alkali-Catalyzed MethanolysisDocument4 pagesProduction of Sunflower Oil Methyl Esters by Optimized Alkali-Catalyzed MethanolysisMáximo Décimo MeridioNo ratings yet
- Chapter 3: Hydrolysis of Vegetable Oils and Animal FatDocument27 pagesChapter 3: Hydrolysis of Vegetable Oils and Animal FatSamiNo ratings yet
- Sythesis of Bioavture Through Hydrodeoxygenation and Catalytic Cracking From Oleic Acid Using Nimo/Zeolit CatalystDocument5 pagesSythesis of Bioavture Through Hydrodeoxygenation and Catalytic Cracking From Oleic Acid Using Nimo/Zeolit Catalystaisyahzafira26No ratings yet
- AasdarticleDocument9 pagesAasdarticleCamilaumNo ratings yet
- 2019 Hura CrepitansDocument8 pages2019 Hura CrepitansNONVIHO GuévaraNo ratings yet
- NMR020008 1 5.0111708Document10 pagesNMR020008 1 5.0111708Sahri YantiNo ratings yet
- I.A Jimoh, Rudyk S.N and Søgaard E.GDocument28 pagesI.A Jimoh, Rudyk S.N and Søgaard E.Graanja2No ratings yet
- Esterification of Oleic Acid To Biodiesel Catalyzed by A Highly Acidic Carbonaceous CatalystDocument10 pagesEsterification of Oleic Acid To Biodiesel Catalyzed by A Highly Acidic Carbonaceous CatalystArif HidayatNo ratings yet
- Kalium HydroxideZirconia Pillared Bentonite For Pa PDFDocument8 pagesKalium HydroxideZirconia Pillared Bentonite For Pa PDFChandra WauranNo ratings yet
- Final ProjectDocument40 pagesFinal Projectmurangiri maronNo ratings yet
- 44 1 Anaheim 03-99 0018Document5 pages44 1 Anaheim 03-99 0018Anca DumitruNo ratings yet
- Date SeedsDocument8 pagesDate SeedsValeria MurilloNo ratings yet
- Hydroprocessing of Palm Oil Using Rh/HZSM-5 For The Production of Biojet Fuel in A Fixed Bed ReactorDocument12 pagesHydroprocessing of Palm Oil Using Rh/HZSM-5 For The Production of Biojet Fuel in A Fixed Bed ReactorNguyễn Văn MinhNo ratings yet
- Production of Biodiesel by Transesterification of Algae Oil With An Assistance of Nano Cao Catalyst Derived From Egg ShellDocument5 pagesProduction of Biodiesel by Transesterification of Algae Oil With An Assistance of Nano Cao Catalyst Derived From Egg ShellMari MuthuNo ratings yet
- Jatropha CurcasDocument1 pageJatropha CurcasAhmad YuhardiNo ratings yet
- Chowdury DKK (2014)Document6 pagesChowdury DKK (2014)Tomy Wijaya PutraNo ratings yet
- Utilization of Waste Freshwater Mussel Shell As An Economic Catalyst For Biodiesel ProductionDocument9 pagesUtilization of Waste Freshwater Mussel Shell As An Economic Catalyst For Biodiesel ProductionDarshak SakhreliyaNo ratings yet
- Biodiesel Production in Nigeria Using Cocoa Pod Ash As A Catalyst BaseDocument4 pagesBiodiesel Production in Nigeria Using Cocoa Pod Ash As A Catalyst BaseInternational Journal of Science and Engineering InvestigationsNo ratings yet
- Spe 179775 MSDocument15 pagesSpe 179775 MSIbrahim SalahudinNo ratings yet
- Bacterial Degradation of Crude Oil by Gravimetric Analysis: Available Online atDocument7 pagesBacterial Degradation of Crude Oil by Gravimetric Analysis: Available Online atSandrianii SaputriiNo ratings yet
- Transesterification Reaction Kinetics of Jatropha Oil For Biodiesel ProductionDocument8 pagesTransesterification Reaction Kinetics of Jatropha Oil For Biodiesel ProductionAmr TarekNo ratings yet
- Methyl Ester Synthesis of Crude Palm Oil Off GradeDocument11 pagesMethyl Ester Synthesis of Crude Palm Oil Off GradeDiar RickiNo ratings yet
- Sust Studies 1Document8 pagesSust Studies 1Mohammad Rakib UddinNo ratings yet
- Akmar 22 1 20Document9 pagesAkmar 22 1 20Becky LawnsNo ratings yet
- Transesterification Reaction Kinetics of Jatropha Oil For Biodiesel ProductionDocument8 pagesTransesterification Reaction Kinetics of Jatropha Oil For Biodiesel ProductionMARTIN PINEDA TAVERANo ratings yet
- 3130-6400-1-RV Edit TEADocument9 pages3130-6400-1-RV Edit TEAMuhammad NopriyansyahNo ratings yet
- Renewable Energy: Jerekias Gandure, Clever Ketlogetswe, Abraham TemuDocument7 pagesRenewable Energy: Jerekias Gandure, Clever Ketlogetswe, Abraham TemuLacramioara Iuliana VladulescuNo ratings yet
- Attacus Ricinii (Eri) Pupae Oil As An Alternative Feedstock For The Production of BiofuelDocument3 pagesAttacus Ricinii (Eri) Pupae Oil As An Alternative Feedstock For The Production of BiofuelWARP-World Academy of Research and PublicationNo ratings yet
- SET REPORT (1) - Converted - CompressedDocument15 pagesSET REPORT (1) - Converted - CompressedvscdNo ratings yet
- Bioresource Technology: Ping Yan, Mang Lu, Qin Yang, Hai-Ling Zhang, Zhong-Zhi Zhang, Rong ChenDocument5 pagesBioresource Technology: Ping Yan, Mang Lu, Qin Yang, Hai-Ling Zhang, Zhong-Zhi Zhang, Rong ChenAkmael Al-hajjNo ratings yet
- Set ReportDocument16 pagesSet ReportvscdNo ratings yet
- IJMEIR Muhammad Fauzi Hartono 04211740000023Document10 pagesIJMEIR Muhammad Fauzi Hartono 04211740000023Fauzi HartonoNo ratings yet
- Biolubricant Synthesis From Waste Cooking Oil Via Enzymatic Hydrolysis Followed by Chemical EsterificationDocument6 pagesBiolubricant Synthesis From Waste Cooking Oil Via Enzymatic Hydrolysis Followed by Chemical EsterificationLeonardo IndraNo ratings yet
- Engineering Journal Biodiesel Production by Using CaO-Al2O3 Nano CatalystDocument7 pagesEngineering Journal Biodiesel Production by Using CaO-Al2O3 Nano CatalystEngineering JournalNo ratings yet
- Particle SizeDocument7 pagesParticle SizeyohannesNo ratings yet
- Environmental Studies On The Microbial Degradation of Oil Hydrocarbons and Its Application in Lebanese Oil Polluted Coastal and Marine EcosystemDocument18 pagesEnvironmental Studies On The Microbial Degradation of Oil Hydrocarbons and Its Application in Lebanese Oil Polluted Coastal and Marine EcosystemAgung Raden MangkubumiNo ratings yet
- Calcium Oxide Catalyst Based On Quail Eggshell For Biodiesel Synthesis From Waste Palm OilDocument10 pagesCalcium Oxide Catalyst Based On Quail Eggshell For Biodiesel Synthesis From Waste Palm OilrisfiNo ratings yet
- Transesterification of Neat and Used Frying Oil: Optimization For Biodiesel ProductionDocument8 pagesTransesterification of Neat and Used Frying Oil: Optimization For Biodiesel ProductionCristiNo ratings yet
- Eng-Investigation On Biofuel Production Using Two-Ayoola AyodejiDocument6 pagesEng-Investigation On Biofuel Production Using Two-Ayoola AyodejiImpact JournalsNo ratings yet
- CURRENT PROBLEMS. Alternative Fuels: Use of Baru Oil (Dipteryx Alata Vog.) TO PRODUCDocument4 pagesCURRENT PROBLEMS. Alternative Fuels: Use of Baru Oil (Dipteryx Alata Vog.) TO PRODUCAntonio Carlos Flash BatistaNo ratings yet
- 1 s2.0 S2666893922000391 MainDocument8 pages1 s2.0 S2666893922000391 Mainmohammed seidNo ratings yet
- Application of Heterogeneous Catalyst of Meti Shells (Batissa Violecea L. Von Lamark 1818) in The Production of Biodiesel Based On Moringa Seed OilDocument7 pagesApplication of Heterogeneous Catalyst of Meti Shells (Batissa Violecea L. Von Lamark 1818) in The Production of Biodiesel Based On Moringa Seed Oilbambang_teknikkimiaNo ratings yet
- Bioresource Technology: Preeti Arora, Dilip R. Ranade, Prashant K. DhakephalkarDocument5 pagesBioresource Technology: Preeti Arora, Dilip R. Ranade, Prashant K. DhakephalkarShafira AdlinaNo ratings yet
- SPE 138135 Research For Microbial Conversion of Residual Oil Into Methane in Depleted Oil Fields To Develop New EOR ProcessDocument10 pagesSPE 138135 Research For Microbial Conversion of Residual Oil Into Methane in Depleted Oil Fields To Develop New EOR ProcessSajad FalahNo ratings yet
- Non-Edible Oil of Cerbera Manghas L. Seed From Seram Island - Maluku As Oil Source of Biodiesel ProductionDocument16 pagesNon-Edible Oil of Cerbera Manghas L. Seed From Seram Island - Maluku As Oil Source of Biodiesel Productioncyndy afrilaNo ratings yet
- Synthesis of Biolubricants From Non Edible OilsDocument5 pagesSynthesis of Biolubricants From Non Edible OilsAnonymous kw8Yrp0R5rNo ratings yet
- Microalgal Lipid Extraction and Evaluation of Single-Step Biodiesel ProductionDocument10 pagesMicroalgal Lipid Extraction and Evaluation of Single-Step Biodiesel ProductionMuhammad AminNo ratings yet
- Production of Biodiesel From Waste Cooking Oil by Transesterification Reaction Using Cao/Natural Zeolite CatalystsDocument7 pagesProduction of Biodiesel From Waste Cooking Oil by Transesterification Reaction Using Cao/Natural Zeolite CatalystsDennis ChanNo ratings yet
- Al Sayegh2015Document11 pagesAl Sayegh2015mehrNo ratings yet
- Bio DegradationDocument8 pagesBio DegradationAtif KhanNo ratings yet
- Single Cell Oils: Microbial and Algal OilsFrom EverandSingle Cell Oils: Microbial and Algal OilsZvi CohenNo ratings yet
- How Does The Jigsaw Technique Work?Document9 pagesHow Does The Jigsaw Technique Work?Italo Aarón Milla LozanoNo ratings yet
- Senior RadiographerDocument1 pageSenior Radiographertafi66No ratings yet
- TM Meeting Roles HBDDocument4 pagesTM Meeting Roles HBDHB DayaratneNo ratings yet
- Drone Spraying BeanDocument4 pagesDrone Spraying BeanFitri AudiaNo ratings yet
- AIM R16 WS02 Rear SpoilerDocument30 pagesAIM R16 WS02 Rear SpoilerAnonymous PJP78mSxNo ratings yet
- Project 4Document6 pagesProject 4Trai TranNo ratings yet
- + 011 Trainees (v4.0 Jan 21)Document11 pages+ 011 Trainees (v4.0 Jan 21)Grace DoucetteNo ratings yet
- A Concept of Pilot Plant For Innovative Production of Ammonium FertilizersDocument6 pagesA Concept of Pilot Plant For Innovative Production of Ammonium FertilizersEndah SaraswatiNo ratings yet
- Spanning Space, Horizontal-Span Building Structures, Wolfgang SchuellerDocument752 pagesSpanning Space, Horizontal-Span Building Structures, Wolfgang Schuellerwolfschueller91% (11)
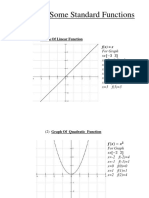
- Lecture 2 Graphs of Some Standard Functions, Shifting of GraphsDocument12 pagesLecture 2 Graphs of Some Standard Functions, Shifting of Graphsghazi membersNo ratings yet
- Corona Rotate Lec MachDocument2 pagesCorona Rotate Lec Machr_chulinNo ratings yet
- Merseyside Risk Assessment V2Document5 pagesMerseyside Risk Assessment V2rob_gorstNo ratings yet
- RElatório Global Teacher 2018 PDFDocument105 pagesRElatório Global Teacher 2018 PDFKimbellyNo ratings yet
- Requerimientos Nutricionales de EquinosDocument112 pagesRequerimientos Nutricionales de EquinosFaby DavalosNo ratings yet
- Astm D2414-21Document9 pagesAstm D2414-21Rajdeep MalNo ratings yet
- The Ecology of DragonsDocument2 pagesThe Ecology of DragonsMarti LelisNo ratings yet
- Book Order FormDocument2 pagesBook Order Formbinaya BehuraNo ratings yet
- How To Get On A TrainDocument4 pagesHow To Get On A TrainreuenNo ratings yet
- 2017 VT CHEM 579 Handout Section 3 (D2L)Document35 pages2017 VT CHEM 579 Handout Section 3 (D2L)Anonymous wPVGEORpjHNo ratings yet
- Iso 11464Document15 pagesIso 11464zvjesosNo ratings yet
- English Language Lab: Sir C R Reddy College of EngineeringDocument76 pagesEnglish Language Lab: Sir C R Reddy College of EngineeringAbhishek BhanNo ratings yet
- Magmatic Garnet in The TriassicDocument27 pagesMagmatic Garnet in The TriassicCARLOS RODRIGUEZNo ratings yet
- DanielsDocument5 pagesDanielstv_whiteboyNo ratings yet
- Holiday HomeworkDocument2 pagesHoliday Homeworkapi-245176244No ratings yet
- KB00194 - DCOM Settings For The OPC Interface That Cause The Following Errors - Advise Returns Error 8004020 - or PIPCDocument1 pageKB00194 - DCOM Settings For The OPC Interface That Cause The Following Errors - Advise Returns Error 8004020 - or PIPCJunior PassosNo ratings yet
























































































Download as pdf or txt
You might also like
- Housing For Swine ProductionDocument87 pagesHousing For Swine ProductionMaria Leonora Theresa100% (3)
- Robert D. Blevins - Flow-Induced Vibration (2001, Krieger Pub Co) PDFDocument254 pagesRobert D. Blevins - Flow-Induced Vibration (2001, Krieger Pub Co) PDFDang Dinh DongNo ratings yet
- Galaxy at War - Star Wars SagaDocument225 pagesGalaxy at War - Star Wars SagaSebastián Rigoni100% (7)
- How To Sell Without Selling - Step - by - Step Marketing Formula To Attract Ready - To - Buy Clients Create Passive Income and Make More Money While Making A DifferenceDocument287 pagesHow To Sell Without Selling - Step - by - Step Marketing Formula To Attract Ready - To - Buy Clients Create Passive Income and Make More Money While Making A Differencedebanikg100% (7)
- Improving Internal Control A Practical Guide For Microfinance InstitutionsDocument83 pagesImproving Internal Control A Practical Guide For Microfinance InstitutionsIon Cara50% (2)
- 2016 Al-Bahry Article - PotentialInHeavyOilBiodegradatDocument13 pages2016 Al-Bahry Article - PotentialInHeavyOilBiodegradatFaqiangNo ratings yet
- Preparation, Characterization and Application of Zeolite-Based Catalyst For Production of Biodiesel From Waste Cooking OilDocument5 pagesPreparation, Characterization and Application of Zeolite-Based Catalyst For Production of Biodiesel From Waste Cooking OilFakhruddin HarahapNo ratings yet
- Determination of Physico-Chemical Properties of Castor Biodiesel A PotentialDocument7 pagesDetermination of Physico-Chemical Properties of Castor Biodiesel A PotentialIAEME PublicationNo ratings yet
- Gassara Et AlDocument6 pagesGassara Et Almushtaq521No ratings yet
- Physico-Chemical Characterization of Algal Oil - Kumar-2011 PDFDocument5 pagesPhysico-Chemical Characterization of Algal Oil - Kumar-2011 PDFrinkanNo ratings yet
- SPE 123303 Bio-Genetic Engineering, Future of Multi Microbial Culture EOR - A Detailed ReportDocument5 pagesSPE 123303 Bio-Genetic Engineering, Future of Multi Microbial Culture EOR - A Detailed ReportSajad FalahNo ratings yet
- Aceite de FrituraDocument8 pagesAceite de FrituraNestor Andres Urbina SuarezNo ratings yet
- Jurnal OkeDocument6 pagesJurnal OkeIlm NafsiNo ratings yet
- 2014 Raed Bacterial Biodegradation of Crude Oil Using LocalDocument8 pages2014 Raed Bacterial Biodegradation of Crude Oil Using LocalFaqiangNo ratings yet
- Production of Sunflower Oil Methyl Esters by Optimized Alkali-Catalyzed MethanolysisDocument4 pagesProduction of Sunflower Oil Methyl Esters by Optimized Alkali-Catalyzed MethanolysisMáximo Décimo MeridioNo ratings yet
- Chapter 3: Hydrolysis of Vegetable Oils and Animal FatDocument27 pagesChapter 3: Hydrolysis of Vegetable Oils and Animal FatSamiNo ratings yet
- Sythesis of Bioavture Through Hydrodeoxygenation and Catalytic Cracking From Oleic Acid Using Nimo/Zeolit CatalystDocument5 pagesSythesis of Bioavture Through Hydrodeoxygenation and Catalytic Cracking From Oleic Acid Using Nimo/Zeolit Catalystaisyahzafira26No ratings yet
- AasdarticleDocument9 pagesAasdarticleCamilaumNo ratings yet
- 2019 Hura CrepitansDocument8 pages2019 Hura CrepitansNONVIHO GuévaraNo ratings yet
- NMR020008 1 5.0111708Document10 pagesNMR020008 1 5.0111708Sahri YantiNo ratings yet
- I.A Jimoh, Rudyk S.N and Søgaard E.GDocument28 pagesI.A Jimoh, Rudyk S.N and Søgaard E.Graanja2No ratings yet
- Esterification of Oleic Acid To Biodiesel Catalyzed by A Highly Acidic Carbonaceous CatalystDocument10 pagesEsterification of Oleic Acid To Biodiesel Catalyzed by A Highly Acidic Carbonaceous CatalystArif HidayatNo ratings yet
- Kalium HydroxideZirconia Pillared Bentonite For Pa PDFDocument8 pagesKalium HydroxideZirconia Pillared Bentonite For Pa PDFChandra WauranNo ratings yet
- Final ProjectDocument40 pagesFinal Projectmurangiri maronNo ratings yet
- 44 1 Anaheim 03-99 0018Document5 pages44 1 Anaheim 03-99 0018Anca DumitruNo ratings yet
- Date SeedsDocument8 pagesDate SeedsValeria MurilloNo ratings yet
- Hydroprocessing of Palm Oil Using Rh/HZSM-5 For The Production of Biojet Fuel in A Fixed Bed ReactorDocument12 pagesHydroprocessing of Palm Oil Using Rh/HZSM-5 For The Production of Biojet Fuel in A Fixed Bed ReactorNguyễn Văn MinhNo ratings yet
- Production of Biodiesel by Transesterification of Algae Oil With An Assistance of Nano Cao Catalyst Derived From Egg ShellDocument5 pagesProduction of Biodiesel by Transesterification of Algae Oil With An Assistance of Nano Cao Catalyst Derived From Egg ShellMari MuthuNo ratings yet
- Jatropha CurcasDocument1 pageJatropha CurcasAhmad YuhardiNo ratings yet
- Chowdury DKK (2014)Document6 pagesChowdury DKK (2014)Tomy Wijaya PutraNo ratings yet
- Utilization of Waste Freshwater Mussel Shell As An Economic Catalyst For Biodiesel ProductionDocument9 pagesUtilization of Waste Freshwater Mussel Shell As An Economic Catalyst For Biodiesel ProductionDarshak SakhreliyaNo ratings yet
- Biodiesel Production in Nigeria Using Cocoa Pod Ash As A Catalyst BaseDocument4 pagesBiodiesel Production in Nigeria Using Cocoa Pod Ash As A Catalyst BaseInternational Journal of Science and Engineering InvestigationsNo ratings yet
- Spe 179775 MSDocument15 pagesSpe 179775 MSIbrahim SalahudinNo ratings yet
- Bacterial Degradation of Crude Oil by Gravimetric Analysis: Available Online atDocument7 pagesBacterial Degradation of Crude Oil by Gravimetric Analysis: Available Online atSandrianii SaputriiNo ratings yet
- Transesterification Reaction Kinetics of Jatropha Oil For Biodiesel ProductionDocument8 pagesTransesterification Reaction Kinetics of Jatropha Oil For Biodiesel ProductionAmr TarekNo ratings yet
- Methyl Ester Synthesis of Crude Palm Oil Off GradeDocument11 pagesMethyl Ester Synthesis of Crude Palm Oil Off GradeDiar RickiNo ratings yet
- Sust Studies 1Document8 pagesSust Studies 1Mohammad Rakib UddinNo ratings yet
- Akmar 22 1 20Document9 pagesAkmar 22 1 20Becky LawnsNo ratings yet
- Transesterification Reaction Kinetics of Jatropha Oil For Biodiesel ProductionDocument8 pagesTransesterification Reaction Kinetics of Jatropha Oil For Biodiesel ProductionMARTIN PINEDA TAVERANo ratings yet
- 3130-6400-1-RV Edit TEADocument9 pages3130-6400-1-RV Edit TEAMuhammad NopriyansyahNo ratings yet
- Renewable Energy: Jerekias Gandure, Clever Ketlogetswe, Abraham TemuDocument7 pagesRenewable Energy: Jerekias Gandure, Clever Ketlogetswe, Abraham TemuLacramioara Iuliana VladulescuNo ratings yet
- Attacus Ricinii (Eri) Pupae Oil As An Alternative Feedstock For The Production of BiofuelDocument3 pagesAttacus Ricinii (Eri) Pupae Oil As An Alternative Feedstock For The Production of BiofuelWARP-World Academy of Research and PublicationNo ratings yet
- SET REPORT (1) - Converted - CompressedDocument15 pagesSET REPORT (1) - Converted - CompressedvscdNo ratings yet
- Bioresource Technology: Ping Yan, Mang Lu, Qin Yang, Hai-Ling Zhang, Zhong-Zhi Zhang, Rong ChenDocument5 pagesBioresource Technology: Ping Yan, Mang Lu, Qin Yang, Hai-Ling Zhang, Zhong-Zhi Zhang, Rong ChenAkmael Al-hajjNo ratings yet
- Set ReportDocument16 pagesSet ReportvscdNo ratings yet
- IJMEIR Muhammad Fauzi Hartono 04211740000023Document10 pagesIJMEIR Muhammad Fauzi Hartono 04211740000023Fauzi HartonoNo ratings yet
- Biolubricant Synthesis From Waste Cooking Oil Via Enzymatic Hydrolysis Followed by Chemical EsterificationDocument6 pagesBiolubricant Synthesis From Waste Cooking Oil Via Enzymatic Hydrolysis Followed by Chemical EsterificationLeonardo IndraNo ratings yet
- Engineering Journal Biodiesel Production by Using CaO-Al2O3 Nano CatalystDocument7 pagesEngineering Journal Biodiesel Production by Using CaO-Al2O3 Nano CatalystEngineering JournalNo ratings yet
- Particle SizeDocument7 pagesParticle SizeyohannesNo ratings yet
- Environmental Studies On The Microbial Degradation of Oil Hydrocarbons and Its Application in Lebanese Oil Polluted Coastal and Marine EcosystemDocument18 pagesEnvironmental Studies On The Microbial Degradation of Oil Hydrocarbons and Its Application in Lebanese Oil Polluted Coastal and Marine EcosystemAgung Raden MangkubumiNo ratings yet
- Calcium Oxide Catalyst Based On Quail Eggshell For Biodiesel Synthesis From Waste Palm OilDocument10 pagesCalcium Oxide Catalyst Based On Quail Eggshell For Biodiesel Synthesis From Waste Palm OilrisfiNo ratings yet
- Transesterification of Neat and Used Frying Oil: Optimization For Biodiesel ProductionDocument8 pagesTransesterification of Neat and Used Frying Oil: Optimization For Biodiesel ProductionCristiNo ratings yet
- Eng-Investigation On Biofuel Production Using Two-Ayoola AyodejiDocument6 pagesEng-Investigation On Biofuel Production Using Two-Ayoola AyodejiImpact JournalsNo ratings yet
- CURRENT PROBLEMS. Alternative Fuels: Use of Baru Oil (Dipteryx Alata Vog.) TO PRODUCDocument4 pagesCURRENT PROBLEMS. Alternative Fuels: Use of Baru Oil (Dipteryx Alata Vog.) TO PRODUCAntonio Carlos Flash BatistaNo ratings yet
- 1 s2.0 S2666893922000391 MainDocument8 pages1 s2.0 S2666893922000391 Mainmohammed seidNo ratings yet
- Application of Heterogeneous Catalyst of Meti Shells (Batissa Violecea L. Von Lamark 1818) in The Production of Biodiesel Based On Moringa Seed OilDocument7 pagesApplication of Heterogeneous Catalyst of Meti Shells (Batissa Violecea L. Von Lamark 1818) in The Production of Biodiesel Based On Moringa Seed Oilbambang_teknikkimiaNo ratings yet
- Bioresource Technology: Preeti Arora, Dilip R. Ranade, Prashant K. DhakephalkarDocument5 pagesBioresource Technology: Preeti Arora, Dilip R. Ranade, Prashant K. DhakephalkarShafira AdlinaNo ratings yet
- SPE 138135 Research For Microbial Conversion of Residual Oil Into Methane in Depleted Oil Fields To Develop New EOR ProcessDocument10 pagesSPE 138135 Research For Microbial Conversion of Residual Oil Into Methane in Depleted Oil Fields To Develop New EOR ProcessSajad FalahNo ratings yet
- Non-Edible Oil of Cerbera Manghas L. Seed From Seram Island - Maluku As Oil Source of Biodiesel ProductionDocument16 pagesNon-Edible Oil of Cerbera Manghas L. Seed From Seram Island - Maluku As Oil Source of Biodiesel Productioncyndy afrilaNo ratings yet
- Synthesis of Biolubricants From Non Edible OilsDocument5 pagesSynthesis of Biolubricants From Non Edible OilsAnonymous kw8Yrp0R5rNo ratings yet
- Microalgal Lipid Extraction and Evaluation of Single-Step Biodiesel ProductionDocument10 pagesMicroalgal Lipid Extraction and Evaluation of Single-Step Biodiesel ProductionMuhammad AminNo ratings yet
- Production of Biodiesel From Waste Cooking Oil by Transesterification Reaction Using Cao/Natural Zeolite CatalystsDocument7 pagesProduction of Biodiesel From Waste Cooking Oil by Transesterification Reaction Using Cao/Natural Zeolite CatalystsDennis ChanNo ratings yet
- Al Sayegh2015Document11 pagesAl Sayegh2015mehrNo ratings yet
- Bio DegradationDocument8 pagesBio DegradationAtif KhanNo ratings yet
- Single Cell Oils: Microbial and Algal OilsFrom EverandSingle Cell Oils: Microbial and Algal OilsZvi CohenNo ratings yet
- How Does The Jigsaw Technique Work?Document9 pagesHow Does The Jigsaw Technique Work?Italo Aarón Milla LozanoNo ratings yet
- Senior RadiographerDocument1 pageSenior Radiographertafi66No ratings yet
- TM Meeting Roles HBDDocument4 pagesTM Meeting Roles HBDHB DayaratneNo ratings yet
- Drone Spraying BeanDocument4 pagesDrone Spraying BeanFitri AudiaNo ratings yet
- AIM R16 WS02 Rear SpoilerDocument30 pagesAIM R16 WS02 Rear SpoilerAnonymous PJP78mSxNo ratings yet
- Project 4Document6 pagesProject 4Trai TranNo ratings yet
- + 011 Trainees (v4.0 Jan 21)Document11 pages+ 011 Trainees (v4.0 Jan 21)Grace DoucetteNo ratings yet
- A Concept of Pilot Plant For Innovative Production of Ammonium FertilizersDocument6 pagesA Concept of Pilot Plant For Innovative Production of Ammonium FertilizersEndah SaraswatiNo ratings yet
- Spanning Space, Horizontal-Span Building Structures, Wolfgang SchuellerDocument752 pagesSpanning Space, Horizontal-Span Building Structures, Wolfgang Schuellerwolfschueller91% (11)
- Lecture 2 Graphs of Some Standard Functions, Shifting of GraphsDocument12 pagesLecture 2 Graphs of Some Standard Functions, Shifting of Graphsghazi membersNo ratings yet
- Corona Rotate Lec MachDocument2 pagesCorona Rotate Lec Machr_chulinNo ratings yet
- Merseyside Risk Assessment V2Document5 pagesMerseyside Risk Assessment V2rob_gorstNo ratings yet
- RElatório Global Teacher 2018 PDFDocument105 pagesRElatório Global Teacher 2018 PDFKimbellyNo ratings yet
- Requerimientos Nutricionales de EquinosDocument112 pagesRequerimientos Nutricionales de EquinosFaby DavalosNo ratings yet
- Astm D2414-21Document9 pagesAstm D2414-21Rajdeep MalNo ratings yet
- The Ecology of DragonsDocument2 pagesThe Ecology of DragonsMarti LelisNo ratings yet
- Book Order FormDocument2 pagesBook Order Formbinaya BehuraNo ratings yet
- How To Get On A TrainDocument4 pagesHow To Get On A TrainreuenNo ratings yet
- 2017 VT CHEM 579 Handout Section 3 (D2L)Document35 pages2017 VT CHEM 579 Handout Section 3 (D2L)Anonymous wPVGEORpjHNo ratings yet
- Iso 11464Document15 pagesIso 11464zvjesosNo ratings yet
- English Language Lab: Sir C R Reddy College of EngineeringDocument76 pagesEnglish Language Lab: Sir C R Reddy College of EngineeringAbhishek BhanNo ratings yet
- Magmatic Garnet in The TriassicDocument27 pagesMagmatic Garnet in The TriassicCARLOS RODRIGUEZNo ratings yet
- DanielsDocument5 pagesDanielstv_whiteboyNo ratings yet
- Holiday HomeworkDocument2 pagesHoliday Homeworkapi-245176244No ratings yet
- KB00194 - DCOM Settings For The OPC Interface That Cause The Following Errors - Advise Returns Error 8004020 - or PIPCDocument1 pageKB00194 - DCOM Settings For The OPC Interface That Cause The Following Errors - Advise Returns Error 8004020 - or PIPCJunior PassosNo ratings yet