Download as pdf or txt
You might also like
- Attention Seeing and Change BlindnessDocument28 pagesAttention Seeing and Change BlindnessGirba EmiliaNo ratings yet
- Exam Practice 1Document1 pageExam Practice 1monicanacaNo ratings yet
- An Accurate Comparison of Methods For Quantifying Variable Importance in Artificial Neural Networks Using Simulated Data-2004Document9 pagesAn Accurate Comparison of Methods For Quantifying Variable Importance in Artificial Neural Networks Using Simulated Data-2004Amine OsmaniNo ratings yet
- Kanban KDocument18 pagesKanban KONE STEP CLOSERNo ratings yet
- Predictive Models in Ecology: Comparison of Performances and Assessment of ApplicabilityDocument17 pagesPredictive Models in Ecology: Comparison of Performances and Assessment of ApplicabilityCatalin CrisanNo ratings yet
- Legendre Fortin MER 2010Document14 pagesLegendre Fortin MER 2010Catalina Verdugo ArriagadaNo ratings yet
- The Impact of Mathematics On Cellular and Molecular BiologyDocument10 pagesThe Impact of Mathematics On Cellular and Molecular Biologydevasena1991No ratings yet
- Checar RedesDocument11 pagesChecar RedesChepis JrfNo ratings yet
- Applications of Graph Theory To Landscape GeneticsDocument11 pagesApplications of Graph Theory To Landscape GeneticsKaritoNo ratings yet
- Re V Iew and Comparison of Methods To STDocument16 pagesRe V Iew and Comparison of Methods To STSaiNo ratings yet
- Applications of Artificial Neural Networks in Foundation EngineeringDocument25 pagesApplications of Artificial Neural Networks in Foundation Engineeringgreenday3100% (1)
- The Appropriate Use of Approximate Entropy and Sample Entropy With Short Data SetsDocument35 pagesThe Appropriate Use of Approximate Entropy and Sample Entropy With Short Data SetskashNo ratings yet
- Porosity and Permeability Estimation Using Neural Network Approach From Well Log DataDocument9 pagesPorosity and Permeability Estimation Using Neural Network Approach From Well Log DataserlyNo ratings yet
- The Study of Periphery Uniqueness and Balance in Ecological NetworksDocument11 pagesThe Study of Periphery Uniqueness and Balance in Ecological Networkschacko4mailNo ratings yet
- 01 de B Harrington 001 JiDocument18 pages01 de B Harrington 001 JiHadjer KermNo ratings yet
- Functional and Effective Connectivity Neuroimaging: A SynthesisDocument23 pagesFunctional and Effective Connectivity Neuroimaging: A SynthesisAnonymous txyvmbYNo ratings yet
- Fuzzy Neural Computing of Coffee and Tainted-Water Data From An Electronic NoseDocument6 pagesFuzzy Neural Computing of Coffee and Tainted-Water Data From An Electronic Noseyohanna maldonadoNo ratings yet
- Ijbc 2010 1Document7 pagesIjbc 2010 1Pedro GarciaNo ratings yet
- Network-Based Regulatory Pathways AnalysisDocument11 pagesNetwork-Based Regulatory Pathways Analysispanna1No ratings yet
- Ecography - 2013 - Merow - A Practical Guide To MaxEnt For Modeling Species Distributions What It Does and Why InputsDocument12 pagesEcography - 2013 - Merow - A Practical Guide To MaxEnt For Modeling Species Distributions What It Does and Why InputsThazin HtayNo ratings yet
- (McRae Et Al, 2008) Using Circuit Theory To Model Connectivity in Ecology, Evolution, and ConservationDocument14 pages(McRae Et Al, 2008) Using Circuit Theory To Model Connectivity in Ecology, Evolution, and ConservationOscar Leonardo Aaron Arizpe VicencioNo ratings yet
- Research 3Document35 pagesResearch 3Prateek PatidarNo ratings yet
- Deterministic Insight Into ANN Model Performance For Storm Runoff SimulationDocument16 pagesDeterministic Insight Into ANN Model Performance For Storm Runoff SimulationDavid JimenezNo ratings yet
- A Practical Guide To Maxent For Modeling Species' Distributions: What It Does, and Why Inputs and Settings MatterDocument12 pagesA Practical Guide To Maxent For Modeling Species' Distributions: What It Does, and Why Inputs and Settings MatterDiego Parra MuñozNo ratings yet
- MANOVA No Parametrico PDFDocument15 pagesMANOVA No Parametrico PDFjesusNo ratings yet
- Genus Statistics of The Virgo N-Body Simulations and The 1.2-Jy Redshift SurveyDocument20 pagesGenus Statistics of The Virgo N-Body Simulations and The 1.2-Jy Redshift Surveyneeraj12121No ratings yet
- Jackson 1993 - Stopping Rules in PCADocument12 pagesJackson 1993 - Stopping Rules in PCAvojarufosiNo ratings yet
- Yang Jian 2007Document10 pagesYang Jian 2007Muhammad KamranNo ratings yet
- Forecasting With Artificial Neural Networks: The State of The ArtDocument29 pagesForecasting With Artificial Neural Networks: The State of The ArtYt YtNo ratings yet
- Permanova PaperDocument15 pagesPermanova PaperjpallotNo ratings yet
- ANDERSON. 2001. A New Method For Non-Parametric Multivariate Analysis of VarianceDocument23 pagesANDERSON. 2001. A New Method For Non-Parametric Multivariate Analysis of VarianceIchtus Soluções AmbientaisNo ratings yet
- A Comparison of Methods For The Statistical Analys PDFDocument24 pagesA Comparison of Methods For The Statistical Analys PDFRiza Wara ElzatiNo ratings yet
- Guia para Modelar EcosistemasDocument13 pagesGuia para Modelar EcosistemasOscar GiovanniNo ratings yet
- Modeling of Species Distributions With Maxent: New Extensions and A Comprehensive EvaluationDocument15 pagesModeling of Species Distributions With Maxent: New Extensions and A Comprehensive EvaluationDavid AarónNo ratings yet
- 007 2020testDocument20 pages007 2020testlinglong.kong2008No ratings yet
- Journal of Biomechanics: W. Sun, J.H. Jin, M.P. Reed, F.S. Gayzik, K.A. Danelson, C.R. Bass, J.Y. Zhang, J.D. RuppDocument8 pagesJournal of Biomechanics: W. Sun, J.H. Jin, M.P. Reed, F.S. Gayzik, K.A. Danelson, C.R. Bass, J.Y. Zhang, J.D. RuppStefanita CiunelNo ratings yet
- Franks Etal 2009 GoG Social NetworksDocument11 pagesFranks Etal 2009 GoG Social NetworksRodrigo PedrosaNo ratings yet
- The Structure of Multi-Neuron Firing Patterns in Primate RetinaDocument13 pagesThe Structure of Multi-Neuron Firing Patterns in Primate RetinaSebastián GallegosNo ratings yet
- Thesis 5 p60-72 CCA New MethodDocument14 pagesThesis 5 p60-72 CCA New MethodLIDIA PUGLIESSANo ratings yet
- BRM C12Document8 pagesBRM C12shubham ghodkeNo ratings yet
- All MAR17Document1,571 pagesAll MAR17mkromelNo ratings yet
- Classifying Soil Structure Using Neural Networks: ElsevierDocument8 pagesClassifying Soil Structure Using Neural Networks: ElseviermanjubondNo ratings yet
- (2016) Analysis of Neuronal Spike Trains DeconstructedDocument39 pages(2016) Analysis of Neuronal Spike Trains DeconstructedDiego Soto ChavezNo ratings yet
- Renner WartonDocument9 pagesRenner WartonMario Alejandro López PérezNo ratings yet
- A Tutorial Review of Functional Connectivity Analysis Methods and Their Interpretational PitfallsDocument23 pagesA Tutorial Review of Functional Connectivity Analysis Methods and Their Interpretational PitfallsBryan ZhengNo ratings yet
- Neural Networks: Arlex Oscar Marín García, Markus Franziskus Müller, Kaspar Schindler, Christian RummelDocument11 pagesNeural Networks: Arlex Oscar Marín García, Markus Franziskus Müller, Kaspar Schindler, Christian RummelAki SunshinNo ratings yet
- Causal Discovery From Temporally Aggregated Time SeriesDocument10 pagesCausal Discovery From Temporally Aggregated Time SeriessahandNo ratings yet
- Data Optimization With Multilayer Perceptron Neural Network and Using New Pattern in Decision Tree ComparativelyDocument8 pagesData Optimization With Multilayer Perceptron Neural Network and Using New Pattern in Decision Tree ComparativelyAkshita dammuNo ratings yet
- A Practical Guide To MaxEntDocument12 pagesA Practical Guide To MaxEntFajar SidiqNo ratings yet
- Carletti 2020 J. Phys. Complex. 1 035006Document17 pagesCarletti 2020 J. Phys. Complex. 1 035006Jose Gregorio Rodriguez VilarrealNo ratings yet
- Using Bayesian Networks To Analyze Expression Data: Nir Friedman Michal LinialDocument25 pagesUsing Bayesian Networks To Analyze Expression Data: Nir Friedman Michal LinialAgung Prihadi SuharyonoNo ratings yet
- Using Bayesian Networks To Analyze Expression DataDocument20 pagesUsing Bayesian Networks To Analyze Expression DataSaad Murtaza KhanNo ratings yet
- 1 s2.0 S089662731931044X MainDocument19 pages1 s2.0 S089662731931044X MainJorge Miranda JaimeNo ratings yet
- Podlaski Machens 2024Document55 pagesPodlaski Machens 2024ckmachensNo ratings yet
- A Comparison of The Mahalanobis-Taguchi System To A Standard Statistical Method For Defect DetectionDocument9 pagesA Comparison of The Mahalanobis-Taguchi System To A Standard Statistical Method For Defect Detection김형진No ratings yet
- Original Contribution: Performance of Ultrasound Echo Decomposition Using Singular Spectrum AnalysisDocument8 pagesOriginal Contribution: Performance of Ultrasound Echo Decomposition Using Singular Spectrum Analysismoonrise1985No ratings yet
- Working Guide To Boosted Regression Trees'Document12 pagesWorking Guide To Boosted Regression Trees'Cyrus RayNo ratings yet
- SC SMDocument26 pagesSC SMJavi ErNo ratings yet
- Tree-Structured Recurrent Switching Linear Dynamical Systems For Multi-Scale ModelingDocument17 pagesTree-Structured Recurrent Switching Linear Dynamical Systems For Multi-Scale ModelingOwenliey LiuNo ratings yet
- sample (1)Document2 pagessample (1)jsyadav.nithNo ratings yet
- Tutorial-05 Effective Stress and CapillarityDocument3 pagesTutorial-05 Effective Stress and Capillarityjsyadav.nithNo ratings yet
- New Microsoft Word DocumentDocument16 pagesNew Microsoft Word Documentjsyadav.nithNo ratings yet
- BrochureDocument2 pagesBrochurejsyadav.nithNo ratings yet
- Unit 3.2 Basic Soil Properties-ISCDocument20 pagesUnit 3.2 Basic Soil Properties-ISCjsyadav.nithNo ratings yet
- Home Assignment-1 SOLUTIONDocument9 pagesHome Assignment-1 SOLUTIONjsyadav.nithNo ratings yet
- Tutorial 07Document5 pagesTutorial 07jsyadav.nithNo ratings yet
- Quillbot Invoice AzZN4UU0g8zVzDVazDocument1 pageQuillbot Invoice AzZN4UU0g8zVzDVazjsyadav.nithNo ratings yet
- Basic Definitions and Relationships - PPTDocument20 pagesBasic Definitions and Relationships - PPTjsyadav.nithNo ratings yet
- Marks Award - Construction Technology and ManagementDocument5 pagesMarks Award - Construction Technology and Managementjsyadav.nithNo ratings yet
- Antivirus Software and Internet Security For Your PC or Mac - McAfeeDocument1 pageAntivirus Software and Internet Security For Your PC or Mac - McAfeejsyadav.nithNo ratings yet
- Determination of Index PropertiesDocument85 pagesDetermination of Index Propertiesjsyadav.nithNo ratings yet
- Question Paper Concrete TechnologyDocument8 pagesQuestion Paper Concrete Technologyjsyadav.nithNo ratings yet
- JnlnewDocument6 pagesJnlnewjsyadav.nithNo ratings yet
- Final Cut List of Internal & External Reappear Examinees For NovDec 2023 Examination Session.Document22 pagesFinal Cut List of Internal & External Reappear Examinees For NovDec 2023 Examination Session.jsyadav.nithNo ratings yet
- Marks Award - Concrete TechnologyDocument5 pagesMarks Award - Concrete Technologyjsyadav.nithNo ratings yet
- Quiz 2Document7 pagesQuiz 2jsyadav.nithNo ratings yet
- 2023 23 AuthorDocument20 pages2023 23 Authorjsyadav.nithNo ratings yet
- Review Comments IGJDocument2 pagesReview Comments IGJjsyadav.nithNo ratings yet
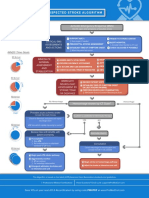
- Suspected Stroke Algorithm: NINDS Time GoalsDocument1 pageSuspected Stroke Algorithm: NINDS Time Goalsjohndoe1995No ratings yet
- All Reflections E001 - E002Document80 pagesAll Reflections E001 - E002PsycOwl ꨄNo ratings yet
- ND - Disturbed Thought ProcessDocument2 pagesND - Disturbed Thought ProcessHu DawiNo ratings yet
- CNS Case PresentationDocument9 pagesCNS Case Presentation073-NAGULAN SHIVAKUMARNo ratings yet
- Written Report Group 3 Ece 16Document4 pagesWritten Report Group 3 Ece 16Jamila PandicalNo ratings yet
- States of ConsciousnessDocument29 pagesStates of ConsciousnessBasel MhaimeedNo ratings yet
- TBI What The Lawyer Needs To Know For Website FINAL Cleaned1Document39 pagesTBI What The Lawyer Needs To Know For Website FINAL Cleaned1bendeni199632No ratings yet
- Lesson 5 Learning Styles and Multiple IntelligencesDocument50 pagesLesson 5 Learning Styles and Multiple IntelligencesOfficial Lara Delos Santos83% (6)
- Mcat NotesDocument2 pagesMcat NotesRiver WalterNo ratings yet
- Conners PresentationDocument22 pagesConners PresentationMrigank Aggarwal100% (1)
- Chapter 15 - Detecting The EnvironmentDocument5 pagesChapter 15 - Detecting The EnvironmentGrace LeungNo ratings yet
- Eye Movement Desensitization and Reprocessing EMDRDocument5 pagesEye Movement Desensitization and Reprocessing EMDRSalem alarjaniNo ratings yet
- Fall 2022 - ZOO403 - 1 - BC190408012Document2 pagesFall 2022 - ZOO403 - 1 - BC190408012Misbah NoreenNo ratings yet
- Study Guide QuestionsDocument4 pagesStudy Guide QuestionszaidaNo ratings yet
- Powerpoint Dan Paul InfancyDocument19 pagesPowerpoint Dan Paul InfancylawNo ratings yet
- Pain Management in Critically Ill Patient: Prof. Dr. Andi Husni Tanra, PHD, SP - An-Kic-KmnDocument42 pagesPain Management in Critically Ill Patient: Prof. Dr. Andi Husni Tanra, PHD, SP - An-Kic-KmnReza Prakosa Sedyatama100% (1)
- Pediatric RehabilitationDocument27 pagesPediatric RehabilitationAstri230% (1)
- Jurnal EEGDocument17 pagesJurnal EEGNurul FajriNo ratings yet
- ResearchDocument4 pagesResearchDave Delos ReyesNo ratings yet
- Mental Status ExaminationDocument7 pagesMental Status ExaminationUdaya Sree100% (4)
- ch15 APR LectureDocument60 pagesch15 APR LectureDrGasnasNo ratings yet
- CASE REPORT PsychiatryDocument6 pagesCASE REPORT PsychiatryTJ LoNo ratings yet
- New PE 1 MODULEDocument56 pagesNew PE 1 MODULEJake GuzmanNo ratings yet
- Psy Chapter 2Document146 pagesPsy Chapter 2OceanNo ratings yet
- Enlarged Peripheral Nerves On LeprosyDocument6 pagesEnlarged Peripheral Nerves On LeprosyGarrett SimpsonNo ratings yet
- Research PaperDocument8 pagesResearch Paperapi-449021066No ratings yet
- RB at and Somatic EducationDocument5 pagesRB at and Somatic Educationdsangax100% (2)
- Study Comparasi Terapi Slow Deep Breathing Dan GuiDocument10 pagesStudy Comparasi Terapi Slow Deep Breathing Dan GuiKimasNo ratings yet