Download as docx, pdf, or txt
You might also like
- Polymer Handbook (4th Edition) - Wiley-Blackwell (1999) PDFDocument2,366 pagesPolymer Handbook (4th Edition) - Wiley-Blackwell (1999) PDFLuis Fernando Quiñónez Cadena67% (3)
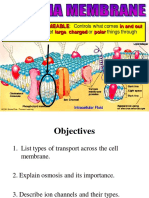
- Transport Across Cell Membranes-1Document7 pagesTransport Across Cell Membranes-1Ifeoma JulietNo ratings yet
- Cell PhysiologyDocument36 pagesCell PhysiologyRayonesh RayanaNo ratings yet
- Animal Physiology Theory 2Document18 pagesAnimal Physiology Theory 2ao868598No ratings yet
- The Permeability of The Plasma MembraneDocument32 pagesThe Permeability of The Plasma Membranenozel77No ratings yet
- 1.1 Transport of SubstancesDocument25 pages1.1 Transport of SubstancesRayonesh RayanaNo ratings yet
- 13-Movement of Substances Across The PDocument8 pages13-Movement of Substances Across The PAbbas TalibNo ratings yet
- Transport Across The Cell Membrane - OdpDocument67 pagesTransport Across The Cell Membrane - OdpSilver CypressNo ratings yet
- Screenshot 2023-02-07 at 9.06.14 PMDocument92 pagesScreenshot 2023-02-07 at 9.06.14 PMMohammed RamzyNo ratings yet
- Transport Across Membranes FinalDocument77 pagesTransport Across Membranes FinalLorraine Eve Abila100% (1)
- TransportDocument65 pagesTransportsiddhi meenaNo ratings yet
- Transport Across The Cell Surface MembraneDocument31 pagesTransport Across The Cell Surface MembraneKalia MckoyNo ratings yet
- Transport Across The Cell Surface MembraneDocument31 pagesTransport Across The Cell Surface MembraneKalia MckoyNo ratings yet
- Gen Bio Transport MechanismDocument46 pagesGen Bio Transport MechanismArianne QuilarioNo ratings yet
- Lect 2 Membrane TransportDocument29 pagesLect 2 Membrane Transportramadan100% (1)
- 2222 TransportDocument42 pages2222 TransportaddwrNo ratings yet
- Cell Transport and HomeostasisDocument2 pagesCell Transport and Homeostasishomamunfat100% (1)
- DiffusionDocument6 pagesDiffusionkrithiga NNo ratings yet
- Drug Absorption-WPS OfficeDocument6 pagesDrug Absorption-WPS Officeloknath sur parthoNo ratings yet
- Cellular Transport ConceptDocument7 pagesCellular Transport ConceptRuhul Qudus NaimNo ratings yet
- The Big Picture: Diffusion: Movement of Particles From A Region of Higher Concentration To A Region of LowDocument5 pagesThe Big Picture: Diffusion: Movement of Particles From A Region of Higher Concentration To A Region of LowNicolas Gomez CombellasNo ratings yet
- Cell Lecture 5-1Document45 pagesCell Lecture 5-1gracechirangoNo ratings yet
- Chapter 1 Doc 4Document4 pagesChapter 1 Doc 4Enju ChipepeNo ratings yet
- Cell Transport MechanismsDocument8 pagesCell Transport MechanismsNhorielyn Joy Disbaro BaltazarNo ratings yet
- CH 3Document37 pagesCH 3Viraat SinghNo ratings yet
- Presentation by AhmadDocument16 pagesPresentation by AhmadAhmad AslamNo ratings yet
- Water MovementDocument13 pagesWater Movementroteromano5No ratings yet
- Transport Across The Cell MembraneDocument16 pagesTransport Across The Cell MembraneJagruti MaratheNo ratings yet
- Membrane Transport ABDocument38 pagesMembrane Transport ABdj77djcqy7No ratings yet
- Passive Transport - WikipediaDocument17 pagesPassive Transport - Wikipediadevotionaldaily4No ratings yet
- Facilitated DiffusionDocument16 pagesFacilitated DiffusionNeveah KayeNo ratings yet
- Cell ExchangesDocument28 pagesCell ExchangesBenzerari ManaNo ratings yet
- Membrane Permiability and Transport: Donna Mae I. Ilar Bs Biology 3ADocument20 pagesMembrane Permiability and Transport: Donna Mae I. Ilar Bs Biology 3ADonna IlarNo ratings yet
- Transport Across MembraneDocument13 pagesTransport Across MembraneHridyanshu Singh RoyNo ratings yet
- Hey GuysDocument2 pagesHey GuysJaidelNo ratings yet
- Document 16Document8 pagesDocument 16Nur HalisahNo ratings yet
- Group 3 Report Physio LecDocument27 pagesGroup 3 Report Physio Lecたこ ゆきNo ratings yet
- Cell Membranes and Transport 2Document49 pagesCell Membranes and Transport 2Rohan PaneruNo ratings yet
- Note For Grade 10Document10 pagesNote For Grade 10peter20000.npkNo ratings yet
- Gene103Lab DiffusionandOsmosisDocument5 pagesGene103Lab DiffusionandOsmosisbodurr571No ratings yet
- Transport of Systems in Microbial CellsDocument25 pagesTransport of Systems in Microbial CellsDiptomoy Bhattacharya100% (1)
- Cell Membrane TransportDocument48 pagesCell Membrane TransportIndra3SHSNo ratings yet
- Chapter 4 Transport Across Cell MembranesDocument10 pagesChapter 4 Transport Across Cell Membranesnoreen dorani100% (1)
- Cellular FunctionsDocument26 pagesCellular FunctionsNILA MAZONNo ratings yet
- Passive Transport: Openstax CollegeDocument8 pagesPassive Transport: Openstax Collegeanon_104510234No ratings yet
- Chapter 5: Homeostasis and Transport Lesson 5.2: Cell Transport-Passive and ActiveDocument10 pagesChapter 5: Homeostasis and Transport Lesson 5.2: Cell Transport-Passive and ActiveVanshika VermaNo ratings yet
- Active Transport: Zed Malenab Kurt Sebastian MagbooDocument16 pagesActive Transport: Zed Malenab Kurt Sebastian MagbooZed MalenabNo ratings yet
- AssignmentDocument6 pagesAssignmentsylvia anneNo ratings yet
- Cell Membrane Membrane: How Substances Get Across The MembraneDocument15 pagesCell Membrane Membrane: How Substances Get Across The MembraneMariane CarandangNo ratings yet
- AS Biology, Cell Membranes and Transport 1Document49 pagesAS Biology, Cell Membranes and Transport 1sachinNo ratings yet
- Cell Membrane FunctionsDocument60 pagesCell Membrane FunctionsKate Clancy NugaoNo ratings yet
- Membrane TransportDocument74 pagesMembrane TransportMiftahul KhairaniNo ratings yet
- A-Level Biology Transport NotesDocument82 pagesA-Level Biology Transport NotesbirizimanabNo ratings yet
- Transport Mechanism MembraneDocument5 pagesTransport Mechanism MembraneJohn Allen GuisdanNo ratings yet
- DiffusionDocument4 pagesDiffusionROBERTO SHERWIN JR TANDASNo ratings yet
- Las-Q3-Wk6.1 2Document12 pagesLas-Q3-Wk6.1 2Jhude JosephNo ratings yet
- Cell and Its FunctionDocument26 pagesCell and Its FunctionMarielle MatunogNo ratings yet
- BS 1Document12 pagesBS 1JAHANZEB NAZIRNo ratings yet
- Passive and Active Cell Transport MechanismsDocument4 pagesPassive and Active Cell Transport MechanismsAlan VargheseNo ratings yet
- R-Sen Lecture 01bDocument57 pagesR-Sen Lecture 01bhimanshu_agraNo ratings yet
- Polymer Nanocomposites Based On Functionalized Carbon NanotubesDocument31 pagesPolymer Nanocomposites Based On Functionalized Carbon NanotubesClaramaria RodriguezNo ratings yet
- Electrolyte Solutions: by Ali Khidher AlobaidyDocument25 pagesElectrolyte Solutions: by Ali Khidher AlobaidyAlwyn James AgcongayNo ratings yet
- AP Chemistry - Classification of Matter Worksheet: Fill in The Blanks in The Table With The Words in The List BelowDocument3 pagesAP Chemistry - Classification of Matter Worksheet: Fill in The Blanks in The Table With The Words in The List Belowalma silvaNo ratings yet
- Blagdon: Air Operated Double Diaphragm PumpDocument2 pagesBlagdon: Air Operated Double Diaphragm PumpMurthy NarayananNo ratings yet
- Cambridge IGCSE: Combined Science 0653/41Document20 pagesCambridge IGCSE: Combined Science 0653/41Thelma MusvotaNo ratings yet
- Nitroglycerine, C3H5N3O9, Is An Explosive Which, On Detonation, Decomposes RapidlyDocument14 pagesNitroglycerine, C3H5N3O9, Is An Explosive Which, On Detonation, Decomposes Rapidlyapi-25909541No ratings yet
- Silver Conductive Paste - 7440-22-4 - Sigma-AldrichDocument8 pagesSilver Conductive Paste - 7440-22-4 - Sigma-AldrichNaseem chNo ratings yet
- Solid Phase MicroextractionDocument16 pagesSolid Phase MicroextractionvivekbhuchemNo ratings yet
- Internship Report in Textile Wet Processing 2023Document6 pagesInternship Report in Textile Wet Processing 2023Uzair AftabNo ratings yet
- Fire Bricks & SandDocument53 pagesFire Bricks & SandMichael MatshonaNo ratings yet
- Chem022 - To Dye ForDocument4 pagesChem022 - To Dye ForPaige DarbonneNo ratings yet
- Table of Common Polyatomic CationsDocument1 pageTable of Common Polyatomic CationsBongiwe NgobeseNo ratings yet
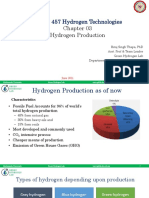
- MEPP 457 Hydrogen TechnologiesDocument88 pagesMEPP 457 Hydrogen TechnologiesAshim LamichhaneNo ratings yet
- Separation TechniquesDocument17 pagesSeparation Techniquesdenise adrieneNo ratings yet
- Chapter 1 Sem 1920Document60 pagesChapter 1 Sem 1920PMNo ratings yet
- NFPA Defining Flammable and Combustible LiquidDocument3 pagesNFPA Defining Flammable and Combustible LiquidkpglemonNo ratings yet
- Hinge Specification GuideDocument4 pagesHinge Specification GuideJoemarie MartinezNo ratings yet
- Accessories For Dosage and Application PDFDocument7 pagesAccessories For Dosage and Application PDFmochkurniawanNo ratings yet
- United States Patent (19) : Obenaus Et Al. (54) Process For Producing High PurityDocument8 pagesUnited States Patent (19) : Obenaus Et Al. (54) Process For Producing High PurityfitriNo ratings yet
- MSDS Colafax 3384Document3 pagesMSDS Colafax 3384mndmattNo ratings yet
- Photoelectrochemical Cells Based On Photosynthetic Systems: A ReviewDocument9 pagesPhotoelectrochemical Cells Based On Photosynthetic Systems: A ReviewMaryam SabouriNo ratings yet
- Calcium Carbonate-Carboxymethyl Chitosan Hybrid Microspheres and Nanospheres For Drug DeliveryDocument6 pagesCalcium Carbonate-Carboxymethyl Chitosan Hybrid Microspheres and Nanospheres For Drug DeliveryPawan Kumar PandeyNo ratings yet
- Chalermpol Innuphat 5610230030 Voltammetric Monitoring of CD (II) by nano-TiO2 Modified Carbon Paste ElectrodeDocument10 pagesChalermpol Innuphat 5610230030 Voltammetric Monitoring of CD (II) by nano-TiO2 Modified Carbon Paste ElectrodewardaninurindahNo ratings yet
- Effect of Chemical and Physical Factors On The Crystallization of Calcium Sulfate in Seawater Reverse Osmosis BrineDocument10 pagesEffect of Chemical and Physical Factors On The Crystallization of Calcium Sulfate in Seawater Reverse Osmosis BrineGilman Ali RezaNo ratings yet
- Inorganic ChemistryDocument2 pagesInorganic ChemistryTaqeeb AbbasNo ratings yet
- Impex Chemicals Product ListDocument11 pagesImpex Chemicals Product Listakartepe1999No ratings yet
- Cemented Carbides 1011Document8 pagesCemented Carbides 1011AB CabralNo ratings yet
- Ease 4 Chemistry Grade 8 19-20Document8 pagesEase 4 Chemistry Grade 8 19-20fajarNo ratings yet
- Shaka Nugusu PDFDocument95 pagesShaka Nugusu PDFEgooNo ratings yet