Download as pdf or txt
You might also like
- Egzamin Teoretyczny Reps Polska Test Z Anatomii I FizjologiiDocument40 pagesEgzamin Teoretyczny Reps Polska Test Z Anatomii I FizjologiiDominika BNo ratings yet
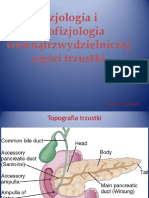
- Trzustka WewnątrzwydzielniczaDocument52 pagesTrzustka WewnątrzwydzielniczaJanNo ratings yet
- Hormony - PytaniaDocument21 pagesHormony - PytaniaAleksandra KNo ratings yet
- Insulina-Kluczowy-Hormon-MetabolicznyDocument8 pagesInsulina-Kluczowy-Hormon-MetabolicznyWiktoria PosłajkoNo ratings yet
- Hormony Peptydowe Wydzialen W Przewodzie PokarmowymDocument3 pagesHormony Peptydowe Wydzialen W Przewodzie PokarmowymAveNooNNo ratings yet
- HormonyDocument13 pagesHormonyJNo ratings yet
- Homeostaza GlukozyDocument27 pagesHomeostaza GlukozyAnna KaczorekNo ratings yet
- BIOCHEMIA-sesja AWFDocument7 pagesBIOCHEMIA-sesja AWFmnik33444No ratings yet
- Funkcje Hormonow CzlowiekaDocument20 pagesFunkcje Hormonow Czlowiekamaciej.majchrowski.szczecinNo ratings yet
- Hormony ŻołądkaDocument18 pagesHormony ŻołądkaKlaudia KorusiewiczNo ratings yet
- Glukoneogeneza I GlikogenolizaDocument17 pagesGlukoneogeneza I Glikogenolizakchowaniec361No ratings yet
- Funkcje Hormonow CzlowiekaDocument17 pagesFunkcje Hormonow CzlowiekaFajna CegłaNo ratings yet
- Kolosy Fizjo 3+4Document6 pagesKolosy Fizjo 3+4m.skwiarniukNo ratings yet
- Glukoneogeneza I GlikogenolizaDocument17 pagesGlukoneogeneza I GlikogenolizaWaifuNoLaifuNo ratings yet
- Farmakologia Hormony 1-50Document8 pagesFarmakologia Hormony 1-50Justyna ZającNo ratings yet
- Biochemia ZadaniaDocument13 pagesBiochemia Zadaniawrobelnatalia781No ratings yet
- (MM2016-1-014) Fizjologia Wysp Trzustkowych I Wydzielania Insuliny: Implikacje Dla InsulinoterapiiDocument11 pages(MM2016-1-014) Fizjologia Wysp Trzustkowych I Wydzielania Insuliny: Implikacje Dla InsulinoterapiiTowarzystwo Edukacji Terapeutycznej100% (1)
- Hormonalna Regulacja Stezenia Glukozy We KrwiDocument21 pagesHormonalna Regulacja Stezenia Glukozy We Krwiwiktoria1805No ratings yet
- Konturek - Wydzielanie WewnętrzneDocument49 pagesKonturek - Wydzielanie Wewnętrznemahoutsukai_monika71% (7)
- 4.8 Inne Procesy MetaboliczneDocument5 pages4.8 Inne Procesy MetabolicznenaczasiebNo ratings yet
- Kolos 4 - WÄ GleDocument56 pagesKolos 4 - WÄ GleOla KoniecznaNo ratings yet
- ĆW 5Document11 pagesĆW 5AXAXNo ratings yet
- Kortyzol - Nadzór Nad Ustrojem W Fizjologii I StresieDocument8 pagesKortyzol - Nadzór Nad Ustrojem W Fizjologii I Stresieccaneccorso1971 ŻukowskaNo ratings yet
- Leki Przysadka Podwzgórze Nadnercza KościDocument59 pagesLeki Przysadka Podwzgórze Nadnercza KościAleksander WarzechaNo ratings yet
- Neurotransmitery, Klasyfikacja, Synteza, Dezaktywacja, Wychwyt - UJDocument74 pagesNeurotransmitery, Klasyfikacja, Synteza, Dezaktywacja, Wychwyt - UJKrzysztof CzaudernaNo ratings yet
- Leczenie CukrzycyDocument5 pagesLeczenie CukrzycyKaja PojdaNo ratings yet
- Kolos 3 HormonyDocument95 pagesKolos 3 HormonyOla KoniecznaNo ratings yet
- Konturek - Układ PokarmowyDocument52 pagesKonturek - Układ Pokarmowymahoutsukai_monika90% (10)
- Aminy-katecholowe-Adrenalina-i-NoradrenalinaDocument8 pagesAminy-katecholowe-Adrenalina-i-NoradrenalinaWiktoria PosłajkoNo ratings yet
- BPZ Egzamin UJ 2Document15 pagesBPZ Egzamin UJ 2jzk34373No ratings yet
- Homeostaza, Krew, Antygeny I PrzeciwciałaDocument14 pagesHomeostaza, Krew, Antygeny I Przeciwciałajagoda.dragonNo ratings yet
- Biologia Układ HormonalnyDocument1 pageBiologia Układ HormonalnyAntek KotarskiNo ratings yet
- Leki HipoglikemizujaceDocument61 pagesLeki HipoglikemizujaceBartek WłodarskiNo ratings yet
- Hormony Rdzenia I Kory NadnerczyDocument19 pagesHormony Rdzenia I Kory NadnerczyArnika WardzyńskaNo ratings yet
- Organizm Człowieka Jako Funkcjonalna Całość-1Document18 pagesOrganizm Człowieka Jako Funkcjonalna Całość-1Oliwier JanoszczykNo ratings yet
- Diabetologia - Prezentacja 2022Document59 pagesDiabetologia - Prezentacja 2022Zuzanna DziwińskaNo ratings yet
- Wykad 3-4Document142 pagesWykad 3-4Gabriela DudekNo ratings yet
- Wykład 10 - Zaburzenia OdżywianiaDocument87 pagesWykład 10 - Zaburzenia OdżywianiaAnka StNo ratings yet
- Odpowiedzi Z BiochemiiDocument34 pagesOdpowiedzi Z Biochemiidominika.cybulska290900No ratings yet
- Fizjologia Dokrewny.Document13 pagesFizjologia Dokrewny.Artur TendelskiNo ratings yet
- Opracowane Zagadnienia BIOCHEMIADocument11 pagesOpracowane Zagadnienia BIOCHEMIAWeronika KozłowskaNo ratings yet
- Żeńskie Hormony PłcioweDocument29 pagesŻeńskie Hormony PłcioweKonrad TheSaintNo ratings yet
- LAKNIENIEDocument16 pagesLAKNIENIEKrisNo ratings yet
- ĆW 3Document4 pagesĆW 3Katarzyna BiegunNo ratings yet
- Markery Zapalenia I Grupy Krwi - SlajdyDocument26 pagesMarkery Zapalenia I Grupy Krwi - Slajdyalina.shot01No ratings yet
- NeuroprzekaźnikiDocument3 pagesNeuroprzekaźnikiOliwia KąpielewskaNo ratings yet
- Diagnostyka Zaburzeń Gospodarki WęglowodanowejDocument12 pagesDiagnostyka Zaburzeń Gospodarki Węglowodanowejg.rainer2001No ratings yet
- Streszcz HormonyDocument35 pagesStreszcz HormonyAnita NowakNo ratings yet
- LIPIDY cz.3 Z Dodatkami (1-80)Document80 pagesLIPIDY cz.3 Z Dodatkami (1-80)Dawid KonieczkoNo ratings yet
- DietetykaDocument16 pagesDietetykaNatalia DargaczNo ratings yet
- Prezentacja 5Document27 pagesPrezentacja 5zosianowak0206No ratings yet
- A.Jeznach Steinhagen-PCOSDocument27 pagesA.Jeznach Steinhagen-PCOSDariaNo ratings yet
- DwunastnicaDocument12 pagesDwunastnicaKrisNo ratings yet
- Hormoni Kore Nadbubrežne Žlezde 1 SASDocument33 pagesHormoni Kore Nadbubrežne Žlezde 1 SASIvana BajunovicNo ratings yet
- Stud MatpptxDocument19 pagesStud MatpptxWeronika KucharskaNo ratings yet
- Cukrzyca - Z RodzinnejDocument86 pagesCukrzyca - Z Rodzinnejc64thorgalNo ratings yet
- 22.cytokiny, Ejkazanoidy, KsenobiotykiDocument63 pages22.cytokiny, Ejkazanoidy, KsenobiotykiankahpfankaNo ratings yet
- Metabolizm Węglowodanów - Utlenianie Glukozy W Warunkach Tlenowych Z Udziałem Np. DrożdżyDocument10 pagesMetabolizm Węglowodanów - Utlenianie Glukozy W Warunkach Tlenowych Z Udziałem Np. DrożdżyNataliaNo ratings yet
- Hormon yDocument1 pageHormon yZannaNo ratings yet
- Neuroetologia BdobreDocument31 pagesNeuroetologia BdobreankahpfankaNo ratings yet
- Neuroetologia AllDocument104 pagesNeuroetologia AllankahpfankaNo ratings yet
- Geometria Płaska (Planimetria) (P. Rozszerzony)Document2 pagesGeometria Płaska (Planimetria) (P. Rozszerzony)ankahpfankaNo ratings yet
- 20 MikroelementyDocument51 pages20 MikroelementyankahpfankaNo ratings yet
- Lista Rankingowa Na Miejsca Szkoleniowe Objęte Rezydenturą - Jesień 2021Document32 pagesLista Rankingowa Na Miejsca Szkoleniowe Objęte Rezydenturą - Jesień 2021ankahpfankaNo ratings yet
- Harmonogram V Roku Lekarskiego Stacjonarnego - Semestr Letni - Rok Akademicki 2023-2024Document21 pagesHarmonogram V Roku Lekarskiego Stacjonarnego - Semestr Letni - Rok Akademicki 2023-2024ankahpfankaNo ratings yet
- PSYCHOPATOLOGIA2Document78 pagesPSYCHOPATOLOGIA2ankahpfankaNo ratings yet
- 19 WitaminyDocument66 pages19 WitaminyankahpfankaNo ratings yet
- AfrodyzjakiDocument8 pagesAfrodyzjakiddadeeeeeNo ratings yet
- POPRAWKA Harciarek ZESTAW PYTAŃDocument9 pagesPOPRAWKA Harciarek ZESTAW PYTAŃkatarzynaanna082No ratings yet
- Hormony. TabelaDocument5 pagesHormony. TabelaAnatolii Ivashko0% (1)
- Nowotwory - Wiadomości Ogólne Ver1Document19 pagesNowotwory - Wiadomości Ogólne Ver1mahoutsukai_monika100% (2)
- Fizjologia Czlowieka Z Elementami Fizjologii Stosowanej I KlinicznejDocument1,170 pagesFizjologia Czlowieka Z Elementami Fizjologii Stosowanej I KlinicznejMadziakNo ratings yet
- Cykl Pracy SercaDocument19 pagesCykl Pracy Sercatinka9179No ratings yet
- Hormony W Sporcie Day One Strenght Performance TrainingDocument72 pagesHormony W Sporcie Day One Strenght Performance TrainingPrzemek RzepaNo ratings yet
- WKB Biologia Konkurs Wymagania 2021 2022Document11 pagesWKB Biologia Konkurs Wymagania 2021 2022HerbatnikNo ratings yet
- Boraks NexusDocument5 pagesBoraks NexusAsia SaraNo ratings yet
- Fizjologia CzowiekaDocument7 pagesFizjologia CzowiekaBaiusiek100% (1)