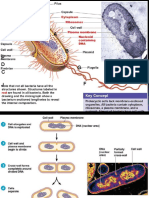
Download as pdf or txt
You might also like
- Examen Traductor Oficial Colombia Modelo1 Ingles EscritoDocument6 pagesExamen Traductor Oficial Colombia Modelo1 Ingles EscritoAlejandro LandinezNo ratings yet
- Gahan 2013 Acidos Nucleicos Efectos InherenciaDocument18 pagesGahan 2013 Acidos Nucleicos Efectos InherenciaKaritoNo ratings yet
- 5 Ipi19356Document9 pages5 Ipi19356Dian Novita AnggraeniNo ratings yet
- 4.牟方详 -EN-Efficacy and safety of recombinant human granulocyte colony-stimulating factor in patients with unexplained recurrent spontaneous abortion:A systematic review and meta-analysis.Document7 pages4.牟方详 -EN-Efficacy and safety of recombinant human granulocyte colony-stimulating factor in patients with unexplained recurrent spontaneous abortion:A systematic review and meta-analysis.johnheshanxiNo ratings yet
- Andrology - 2018 - Panner Selvam - Treatment of Semen Samples With Chymotrypsin Alters The Expression Pattern of SpermDocument6 pagesAndrology - 2018 - Panner Selvam - Treatment of Semen Samples With Chymotrypsin Alters The Expression Pattern of SpermStevenNo ratings yet
- 2008 - Juan Felipe Velez de La Calle - Spermdeoxyribonucleicacidfragmentationasassessedby (Retrieved 2019-04-20)Document8 pages2008 - Juan Felipe Velez de La Calle - Spermdeoxyribonucleicacidfragmentationasassessedby (Retrieved 2019-04-20)Tiffany LamNo ratings yet
- NBT Multi Centric TrialsDocument9 pagesNBT Multi Centric TrialsRakesh RamananNo ratings yet
- A Genetic Mosaic Screen Reveals Ecdysone-Responsive Genes Regulating Drosophila OogenesisDocument15 pagesA Genetic Mosaic Screen Reveals Ecdysone-Responsive Genes Regulating Drosophila Oogenesis奕蓁陳No ratings yet
- Epigenetic Regulation in PlantsDocument33 pagesEpigenetic Regulation in PlantsNina ReisNo ratings yet
- Tempo and Mode of Genome PDFDocument30 pagesTempo and Mode of Genome PDFGinna SolarteNo ratings yet
- 10 1016@j Cell 2016 03 034Document15 pages10 1016@j Cell 2016 03 034Yamile A Rodríguez RiascosNo ratings yet
- A Genomic Diagnostic Tool For Human Endometrial Receptivity Based On The Transcriptomic SignatureDocument26 pagesA Genomic Diagnostic Tool For Human Endometrial Receptivity Based On The Transcriptomic Signatureraulcl1994No ratings yet
- Infection, Genetics and Evolution: Haemaphysalis QinghaiensisDocument15 pagesInfection, Genetics and Evolution: Haemaphysalis QinghaiensisChickencito PerezNo ratings yet
- Analysis of Streptococcal CRISPRs From Human Saliva Reveals Substantial Sequence Diversity Within and Between Subjects Over TimeDocument12 pagesAnalysis of Streptococcal CRISPRs From Human Saliva Reveals Substantial Sequence Diversity Within and Between Subjects Over Timeabcder1234No ratings yet
- C Elegans DissertationDocument7 pagesC Elegans DissertationBuySchoolPapersOnlineSingapore100% (1)
- In Vitro Post-Meiotic Germ Cell Development From Human Embryonic Stem CellsDocument10 pagesIn Vitro Post-Meiotic Germ Cell Development From Human Embryonic Stem Cellsucich23No ratings yet
- Arthritis Rheumatology - 2022 - Hou - Integrative Functional Genomics Identifies Systemic Lupus Erythematosus CausalDocument12 pagesArthritis Rheumatology - 2022 - Hou - Integrative Functional Genomics Identifies Systemic Lupus Erythematosus CausalMaria Alejandra MayaNo ratings yet
- Tissue-Specific Selection of Stable Reference Genes For Real-Time PCR Normalization in An Obese Rat ModelDocument10 pagesTissue-Specific Selection of Stable Reference Genes For Real-Time PCR Normalization in An Obese Rat ModelMohammed AlshinqityNo ratings yet
- 14 ArapDocument8 pages14 ArapThien LuNo ratings yet
- Lactose Persistence Nihms778525Document26 pagesLactose Persistence Nihms778525kaleb16_2No ratings yet
- PIIS001502821400363XDocument7 pagesPIIS001502821400363XzhuzhendongNo ratings yet
- Comparative Biochemistry and Physiology Part A: Molecular & Integrative PhysiologyDocument9 pagesComparative Biochemistry and Physiology Part A: Molecular & Integrative PhysiologyАлександр КолядаNo ratings yet
- Fertility 2021 Abstract-BookDocument119 pagesFertility 2021 Abstract-BookAhmed GhanimNo ratings yet
- Early Development Exposure To Pentachlorophenol Causes Alterations On mRNA Expressions of Caspase Protease Family in Zebrafish EmbryosDocument8 pagesEarly Development Exposure To Pentachlorophenol Causes Alterations On mRNA Expressions of Caspase Protease Family in Zebrafish EmbryosAna Maria RomeroNo ratings yet
- Genetic Signatures of Evolution of The Pluripotency Gene Regulating Network Across MammalsDocument13 pagesGenetic Signatures of Evolution of The Pluripotency Gene Regulating Network Across MammalsSevim AkçağlarNo ratings yet
- Differential Expression of LNC RNADocument12 pagesDifferential Expression of LNC RNAnikhita.chirraNo ratings yet
- Journal of Reproductive Immunology: A 1 A 1 A A B C DDocument4 pagesJournal of Reproductive Immunology: A 1 A 1 A A B C DHaribabu ArumugamNo ratings yet
- Schistosoma ActualidadDocument5 pagesSchistosoma ActualidadWillian EspinozaNo ratings yet
- Nih Public Access: The Genotype-Tissue Expression (Gtex) ProjectDocument13 pagesNih Public Access: The Genotype-Tissue Expression (Gtex) ProjectSandra Milena Sanabria BarreraNo ratings yet
- Pattern of Change in Histone 3 Lysine 9 Acetylation and Histone Deacetylases in Development of Zebrafish EmbryoDocument6 pagesPattern of Change in Histone 3 Lysine 9 Acetylation and Histone Deacetylases in Development of Zebrafish EmbryoTiago TorresNo ratings yet
- Genome Res.-2011-Spencer-325-41Document19 pagesGenome Res.-2011-Spencer-325-41John MistryNo ratings yet
- Penetraciion Espermatica en AlpacasDocument9 pagesPenetraciion Espermatica en AlpacasDaniel Gandarillas EspezuaNo ratings yet
- Paternally Induced Transgenerational Environmental Reprogramming of Metabolic Gene Expression in MammalsDocument13 pagesPaternally Induced Transgenerational Environmental Reprogramming of Metabolic Gene Expression in MammalsAshok KumarNo ratings yet
- CircRNA Expression Profile and Functional Analysis in Testicular Tissue of Patients With Non-Obstructive AzoospermiaDocument10 pagesCircRNA Expression Profile and Functional Analysis in Testicular Tissue of Patients With Non-Obstructive AzoospermiaBahlibiNo ratings yet
- Stanton 2003Document7 pagesStanton 2003Pedro (Official Pedro)No ratings yet
- Vertical Transmissibility of Small RuminantDocument12 pagesVertical Transmissibility of Small RuminantJosé Hiram Sánchez GascaNo ratings yet
- Genome-Wide Identification of RETINOBLASTOMA RELATED 1 Binding Sites in RegulatorsDocument35 pagesGenome-Wide Identification of RETINOBLASTOMA RELATED 1 Binding Sites in RegulatorsletsgopartaeNo ratings yet
- AxolotlDocument12 pagesAxolotlfrancescoNo ratings yet
- Novel Genes and Mutations in Patients Affected by Recurrent Pregnancy LossDocument14 pagesNovel Genes and Mutations in Patients Affected by Recurrent Pregnancy LossarusezNo ratings yet
- iTRAQ and PRM-based Quantitative Proteomics in Early Recurrent Spontaneous Abortion: Biomarkers DiscoveryDocument15 pagesiTRAQ and PRM-based Quantitative Proteomics in Early Recurrent Spontaneous Abortion: Biomarkers DiscoverypriyaNo ratings yet
- Comparative Biochemistry and Physiology, Part C: Linda Bingsohn, Eileen Knorr, Andreas VilcinskasDocument8 pagesComparative Biochemistry and Physiology, Part C: Linda Bingsohn, Eileen Knorr, Andreas VilcinskasbiancacuritibaNo ratings yet
- Garud 2019 PLOSBio EvDynamicsGutMicrobiomeDocument29 pagesGarud 2019 PLOSBio EvDynamicsGutMicrobiomeAnderson MNo ratings yet
- Six2 Regulates Pax9 Expression, Palatogenesis and Craniofacial Bone FormationDocument11 pagesSix2 Regulates Pax9 Expression, Palatogenesis and Craniofacial Bone FormationYerly Ramirez MuñozNo ratings yet
- Biomol NowDocument9 pagesBiomol NowHendra Bayu KurniawanNo ratings yet
- Prenatal Ethanol Exposure and NeocorticalDocument14 pagesPrenatal Ethanol Exposure and Neocorticalpsic.tatianebezerraNo ratings yet
- Timing, Rates and Spectra of Human Germline Mutation: ArticlesDocument11 pagesTiming, Rates and Spectra of Human Germline Mutation: ArticlesAnonymous n2DPWfNuNo ratings yet
- DNA Repair and Replication Links To Pluripotency and Differentiation Capacity of Pig iPS CellsDocument22 pagesDNA Repair and Replication Links To Pluripotency and Differentiation Capacity of Pig iPS CellsmustikaNo ratings yet
- Gordon 2021Document25 pagesGordon 2021ShagorNo ratings yet
- Volume - 41 - 4 - 97 - 3 - MacacaDocument16 pagesVolume - 41 - 4 - 97 - 3 - MacacaAnindyaMustikaNo ratings yet
- Crisp1 and Alopecia Areata in C3H/Hej Mice: Crisp1/Crisp1, Cysteine-Rich Secretory Protein 1, Gene/Protein Crisp1Document12 pagesCrisp1 and Alopecia Areata in C3H/Hej Mice: Crisp1/Crisp1, Cysteine-Rich Secretory Protein 1, Gene/Protein Crisp1Saifuddin HaswareNo ratings yet
- Pi Is 0022534715046030Document8 pagesPi Is 0022534715046030rodesa barnawi17211994No ratings yet
- Abuaish 2021 The Role of Sex Differentiated Variations in Stress Hormones Antiox Neuroimmun Res in Rel To Soc Int Impair in Rodent Model of AutismDocument11 pagesAbuaish 2021 The Role of Sex Differentiated Variations in Stress Hormones Antiox Neuroimmun Res in Rel To Soc Int Impair in Rodent Model of AutismJortegloriaNo ratings yet
- PNAS 2012 Wang 1119459109Document6 pagesPNAS 2012 Wang 1119459109astazangastaNo ratings yet
- Environmental Exposure of Sperm Sex-Chromosomes: A Gender Selection TechniqueDocument9 pagesEnvironmental Exposure of Sperm Sex-Chromosomes: A Gender Selection TechniqueKurniasaryRerungAlloNo ratings yet
- IJKD - News Letter1 20 26Document8 pagesIJKD - News Letter1 20 26Kapil SejpalNo ratings yet
- Shimizu2020 tlr21Document9 pagesShimizu2020 tlr21misilpiyapiyoNo ratings yet
- Analyses of circRNAs Profiles of The Lactating and Nonlactating Crops in PigeonDocument10 pagesAnalyses of circRNAs Profiles of The Lactating and Nonlactating Crops in PigeonBahlibiNo ratings yet
- Association Between Single Nucleotide Polymorphisms Andsexual Precocity in Nellore HeifersDocument9 pagesAssociation Between Single Nucleotide Polymorphisms Andsexual Precocity in Nellore HeifersInaê RegatieriNo ratings yet
- Aneuploidy ConcordanceDocument12 pagesAneuploidy ConcordancejuanNo ratings yet
- 1-s2.0-S004101011730301X-Eric David Merkley Castor Seeds 2017Document41 pages1-s2.0-S004101011730301X-Eric David Merkley Castor Seeds 2017rahi ahmedNo ratings yet
- Summary of The Gene: by Siddhartha Mukherjee | Includes AnalysisFrom EverandSummary of The Gene: by Siddhartha Mukherjee | Includes AnalysisNo ratings yet
- Individual Development & IdentityDocument13 pagesIndividual Development & IdentityTHEA LAURENE SUBANo ratings yet
- Ovary and Embryo CultureDocument4 pagesOvary and Embryo CultureAnandKuttiyanNo ratings yet
- Sex Linkage Worksheet 1: Name Date PeriodDocument2 pagesSex Linkage Worksheet 1: Name Date PeriodAddy Milligan100% (1)
- Bio 131 Cell BioDocument65 pagesBio 131 Cell BioElsie GalvezNo ratings yet
- Practice Test in Chromosome Structure - Question 6: MatchingDocument3 pagesPractice Test in Chromosome Structure - Question 6: MatchingRip GrammarNo ratings yet
- Vertebrate Embryogenesis - Embryological, Cellular, and Genetic Methods (PDFDrive)Document617 pagesVertebrate Embryogenesis - Embryological, Cellular, and Genetic Methods (PDFDrive)AndresNo ratings yet
- DNA DR In-Class ActivitiesDocument15 pagesDNA DR In-Class ActivitiesCéline Engels100% (1)
- Gen - Bio 1 Final ExaminationDocument3 pagesGen - Bio 1 Final ExaminationGrace LimatoNo ratings yet
- Theories of Crime Early Biological TheoriesDocument6 pagesTheories of Crime Early Biological TheoriesCharmis TubilNo ratings yet
- JumpChain Bestowable PowersAbilitiesDocument3 pagesJumpChain Bestowable PowersAbilitiesZiggurathNo ratings yet
- SECOND Semester, AY 2022-2023: Mission VisionDocument34 pagesSECOND Semester, AY 2022-2023: Mission Visionjeyyy BonesssNo ratings yet
- Uhs Mbbs 2Nd Professional Examination Table of Specification For Anatomy (Mcqs & Seqs)Document3 pagesUhs Mbbs 2Nd Professional Examination Table of Specification For Anatomy (Mcqs & Seqs)Farah Khan100% (1)
- A.8. Rule Systems in Chomskyan Theories of LanguageDocument7 pagesA.8. Rule Systems in Chomskyan Theories of LanguageNguyen HalohaloNo ratings yet
- The Human Genome - FinalDocument27 pagesThe Human Genome - FinalAbhi SachdevNo ratings yet
- Biotech-Genetic Sci. TimelineDocument4 pagesBiotech-Genetic Sci. TimelinedreyyNo ratings yet
- GENERAL BIOLOGY ReviewerDocument27 pagesGENERAL BIOLOGY ReviewerBeatriz gannabanNo ratings yet
- A Level Biology Topic 1 AssessmentDocument15 pagesA Level Biology Topic 1 AssessmentgsapkaiteNo ratings yet
- DNH Qualicon Brochure Ribo 1A-0771 1212 WEB PDFDocument11 pagesDNH Qualicon Brochure Ribo 1A-0771 1212 WEB PDFpatricioNo ratings yet
- Vadim Fong - Psychology 12 1b ApproachesDocument2 pagesVadim Fong - Psychology 12 1b ApproachesVadim FongNo ratings yet
- Cbjesccq 09Document24 pagesCbjesccq 09Vinita RamtekeNo ratings yet
- Development TasksDocument4 pagesDevelopment TasksgretutegeruteNo ratings yet
- Lily: Lilium Hybrids: January 2006Document21 pagesLily: Lilium Hybrids: January 2006erikaNo ratings yet
- Bio PBL F5 - HerbariumDocument14 pagesBio PBL F5 - HerbariumYu Han Ong100% (1)
- Behavior GeneticsDocument3 pagesBehavior GeneticsNazreen S MNo ratings yet
- Sc9 PAT Study GuideDocument77 pagesSc9 PAT Study GuideRaymond ORR100% (1)
- Diagnosis: Spinal AtrophyDocument2 pagesDiagnosis: Spinal AtrophyJoelis VerdingasNo ratings yet
- Cultures of EugenicsDocument28 pagesCultures of Eugenicsriichar ramosNo ratings yet
- Bifunctional Polynucleotide Phosphatase/kinase Is An EnzymeDocument10 pagesBifunctional Polynucleotide Phosphatase/kinase Is An Enzymefelipe smithNo ratings yet
- Sci 313.313L - Genetics-Learning Material - Vol. IDocument55 pagesSci 313.313L - Genetics-Learning Material - Vol. IRoselyn BenavidezNo ratings yet