
Teaching Books, Speaking Games
Teaching Books, Speaking Games
You might also like
- Comparative Vertebrate AnatomyDocument11 pagesComparative Vertebrate AnatomyKean GB60% (5)
- Kiss NotesDocument38 pagesKiss NotesNalini Prakash100% (2)
- Textbook of Wild and Zoo AnimalsDocument247 pagesTextbook of Wild and Zoo AnimalsToha Putra100% (1)
- Chapter 1 - History of Zebrafish Resea - 2020 - The Zebrafish in Biomedical Rese PDFDocument12 pagesChapter 1 - History of Zebrafish Resea - 2020 - The Zebrafish in Biomedical Rese PDFNicolas BaronNo ratings yet
- 2012cooper Et Al Wires Development of Bat FlightDocument7 pages2012cooper Et Al Wires Development of Bat Flightapi-237346249No ratings yet
- Developmental Dynamics - 2013 - Sheng - Day 1 Chick DevelopmentDocument11 pagesDevelopmental Dynamics - 2013 - Sheng - Day 1 Chick DevelopmentYasir Iqbal KhanNo ratings yet
- Hendler 1975Document25 pagesHendler 1975byhz7wwdm4No ratings yet
- Integumentary SystemDocument51 pagesIntegumentary Systemmicklemagdy50No ratings yet
- Crustacean Larvae - Academic ScriptDocument12 pagesCrustacean Larvae - Academic ScriptAfridi ShaikhNo ratings yet
- Crustaceae Larval Forms PDFDocument12 pagesCrustaceae Larval Forms PDFAfridi ShaikhNo ratings yet
- Dvdy24087 5 Final As PublishedDocument12 pagesDvdy24087 5 Final As PublishedTahseen FatimaNo ratings yet
- Museum of Comparative Zoology, Harvard University, Cambridge, Massachusetts 02138Document12 pagesMuseum of Comparative Zoology, Harvard University, Cambridge, Massachusetts 02138Julio César Chávez GalarzaNo ratings yet
- Lesson 5 Birth Growth and Reproduction V5Document8 pagesLesson 5 Birth Growth and Reproduction V5JuanNo ratings yet
- Lab Act 1Document4 pagesLab Act 1Ostan Balanquit TeresaNo ratings yet
- Perspectives On The Use of The Baboon in Embryology and Teratology ResearchDocument18 pagesPerspectives On The Use of The Baboon in Embryology and Teratology ResearchYusraNo ratings yet
- Evolution: What Darwin Did Not Know by Then..! [And the Origin of Species Through Species-Branding]From EverandEvolution: What Darwin Did Not Know by Then..! [And the Origin of Species Through Species-Branding]No ratings yet
- 3rd Laboratory Activity AnuraDocument7 pages3rd Laboratory Activity AnuraZia Ammarah SaripNo ratings yet
- Elife 43828Document19 pagesElife 43828jaypasco79No ratings yet
- Zelfstudie Van Blastula Naar 3 KiemlagenDocument18 pagesZelfstudie Van Blastula Naar 3 KiemlagenCheyenne ProvoostNo ratings yet
- Medical Journal of Cell Biology Drosophila Melanogaster Research History, Breakthrough and PerspectivesDocument4 pagesMedical Journal of Cell Biology Drosophila Melanogaster Research History, Breakthrough and PerspectivesIgor DemićNo ratings yet
- Carril Et Al-2015-The Anatomical RecordDocument12 pagesCarril Et Al-2015-The Anatomical RecordrefestufuNo ratings yet
- Burung Matin PDFDocument2 pagesBurung Matin PDFRiskian Alfaro BabtistaNo ratings yet
- Altig & McDiarmid (2007) Morphological Diversity and Evolution of Egg and Clutch Structure in AmphibiansDocument33 pagesAltig & McDiarmid (2007) Morphological Diversity and Evolution of Egg and Clutch Structure in AmphibiansJuan Carlos SánchezNo ratings yet
- Oogenesis and Ovarian Histology of The South American Lizard Salvator MerianaeDocument14 pagesOogenesis and Ovarian Histology of The South American Lizard Salvator MerianaeValeria Garcia ValdezNo ratings yet
- Worksheet Q3 Week 5Document2 pagesWorksheet Q3 Week 5Jaybie TejadaNo ratings yet
- Prototheria: ClassificationDocument5 pagesPrototheria: ClassificationAalamgeer AnsariNo ratings yet
- Anatomy and Development of Oral Cavity and PharynxDocument14 pagesAnatomy and Development of Oral Cavity and PharynxGabriela MacoveanuNo ratings yet
- Evidence of Evolution - Module 6Document35 pagesEvidence of Evolution - Module 6Reign PabloNo ratings yet
- Matthews 1998Document9 pagesMatthews 1998osteopedcursoNo ratings yet
- The History of Vaginal BirthDocument11 pagesThe History of Vaginal BirthdardonnnNo ratings yet
- Evidence of EvolutionDocument15 pagesEvidence of Evolutionsurajkeshari9826No ratings yet
- Evolution of The Mammalian Neck From Developmental, Morpho-Functional, and Paleontological PerspectivesDocument11 pagesEvolution of The Mammalian Neck From Developmental, Morpho-Functional, and Paleontological PerspectivesKo KaNo ratings yet
- Embryologydevelopmental AnatomyDocument62 pagesEmbryologydevelopmental AnatomygirishNo ratings yet
- The Origin of The Turtle Body Plan EvideDocument19 pagesThe Origin of The Turtle Body Plan EvideXweuis Hekuos KweNo ratings yet
- Comparative Anatomy, Embryology & Vestigial StructuresDocument11 pagesComparative Anatomy, Embryology & Vestigial StructuresDaniel BerryNo ratings yet
- Basic Body Plan of Young Mammalian EmbryosDocument57 pagesBasic Body Plan of Young Mammalian EmbryosWin DierNo ratings yet
- Olfaction, Navigation, and The Origin of Isocortex: Francisco Aboitiz and Juan F. MontielDocument12 pagesOlfaction, Navigation, and The Origin of Isocortex: Francisco Aboitiz and Juan F. MontielmioclaenusNo ratings yet
- Blueprint of LifeDocument42 pagesBlueprint of Lifesophiehee1234No ratings yet
- Evidence of Evolution FinalDocument82 pagesEvidence of Evolution FinalCheskaNo ratings yet
- The Phylum CtenophoraDocument3 pagesThe Phylum CtenophoraTI Journals PublishingNo ratings yet
- Development of The Human Tail Bud and Splanchnic Mesenchyme: Original ArticleDocument7 pagesDevelopment of The Human Tail Bud and Splanchnic Mesenchyme: Original Articleproject-247758No ratings yet
- ZootaxaDocument17 pagesZootaxaoliviamuliawatideviNo ratings yet
- Group 4 Teacher'sDocument4 pagesGroup 4 Teacher'sAdrian GabrielNo ratings yet
- Regional Specification in The Early Embryo of The Brittle Star Ophiopholis AculeataDocument16 pagesRegional Specification in The Early Embryo of The Brittle Star Ophiopholis AculeataJoElunico QuezadaNo ratings yet
- Comisuritis Posterior 2Document8 pagesComisuritis Posterior 2VORTIZNo ratings yet
- Gastrulation in The Mouse Embryo: Ultrastructural and Molecular Aspects of Germ Layer MorphogenesisDocument28 pagesGastrulation in The Mouse Embryo: Ultrastructural and Molecular Aspects of Germ Layer MorphogenesisDIEGO FERNANDO URIBE JARAMILLONo ratings yet
- Order Myctophiformes: Blackchins and Lanternfishes by H. Geoffrey Moser and William WatsonDocument49 pagesOrder Myctophiformes: Blackchins and Lanternfishes by H. Geoffrey Moser and William WatsonAcuario Hatchery Los VilosNo ratings yet
- j.1463 6395.2008.00349.x20170103 12513 B3dicm With Cover Page v2Document12 pagesj.1463 6395.2008.00349.x20170103 12513 B3dicm With Cover Page v2Zulema EmirNo ratings yet
- Hendler 1986Document8 pagesHendler 1986byhz7wwdm4No ratings yet
- IESO Preparation Material (Paleontology)Document25 pagesIESO Preparation Material (Paleontology)Science Olympiad Blog67% (3)
- Larva of CrustaceanDocument9 pagesLarva of Crustaceanblloveraastha111No ratings yet
- Larva: Evolutionary TheoryDocument2 pagesLarva: Evolutionary TheoryEdron Bullo JrNo ratings yet
- Lecture 10-Coelomate BauplanDocument59 pagesLecture 10-Coelomate BauplanKago NgomaNo ratings yet
- Seijjjj 378777y36262626363. 4444 22373737374 A - U8ujb Vjhdjdjd. Bbwqwqwoo Rchins - As - Model - System - For - Studying - DevelopmentDocument7 pagesSeijjjj 378777y36262626363. 4444 22373737374 A - U8ujb Vjhdjdjd. Bbwqwqwoo Rchins - As - Model - System - For - Studying - DevelopmentRed DiggerNo ratings yet
- TEAM A BIo 180 Lab Learning Contract 1 Evidence of EvolutionDocument20 pagesTEAM A BIo 180 Lab Learning Contract 1 Evidence of EvolutionJames Carbonell Dela PeñaNo ratings yet
- Sed 202 Biological Science Research Tasks BDocument8 pagesSed 202 Biological Science Research Tasks BBryllie BautistaNo ratings yet
- Fullregenerationoftribasal Polypterusfin PNASDocument9 pagesFullregenerationoftribasal Polypterusfin PNASBagas Lantip PrakasaNo ratings yet
- GASTRULATION-ACTIVITY - DEV-BIO - Bio 102Document6 pagesGASTRULATION-ACTIVITY - DEV-BIO - Bio 102Quentin LopezNo ratings yet
- Donoghue Et Al-2014-PalaeontologyDocument16 pagesDonoghue Et Al-2014-PalaeontologyzhaNo ratings yet
- Introduction To The Zebrafish: MaterialsDocument9 pagesIntroduction To The Zebrafish: MaterialsDogNo ratings yet
- Pakistan Vertebrate FaunaDocument9 pagesPakistan Vertebrate FaunaNosheen MalikNo ratings yet
- Kingdom: Animals: Domain EukaryaDocument25 pagesKingdom: Animals: Domain EukaryaDoods GaldoNo ratings yet
- RTP Las English 5 Q3 Melc 2C - Summarizing Classification Text TypeDocument10 pagesRTP Las English 5 Q3 Melc 2C - Summarizing Classification Text TypePinaka ShungaNo ratings yet
- Kapocs Australia 5-6 PDFDocument21 pagesKapocs Australia 5-6 PDFMohamed HassanNo ratings yet
- Animal KingdomDocument67 pagesAnimal KingdomNaveen Krishna M.KNo ratings yet
- SDADASDASDASDDocument16 pagesSDADASDASDASDAngelaNo ratings yet
- A Manual of Mammalogy With Keys To Families of The World 3rd Edition Ebook PDFDocument62 pagesA Manual of Mammalogy With Keys To Families of The World 3rd Edition Ebook PDFandrea.davis76598% (46)
- 1 Intro To Exotics 7-2023Document56 pages1 Intro To Exotics 7-2023Hailey WilliamsNo ratings yet
- Origin of MammalsDocument26 pagesOrigin of Mammalsjai kumarNo ratings yet
- p.6 Paramount Integrated Science Notes Term 1Document152 pagesp.6 Paramount Integrated Science Notes Term 1Edward Biryetega100% (2)
- Interactive Textbook ScienceDocument464 pagesInteractive Textbook ScienceJose Manuel Rodriguez Cortes100% (3)
- 109Document14 pages109manhtuan15aNo ratings yet
- Britannica Illustrated Science Library - Mammals PDFDocument56 pagesBritannica Illustrated Science Library - Mammals PDFCupitu MadalinaNo ratings yet
- Curious Creatures in Zoology by John Ashton - PDF RoomDocument112 pagesCurious Creatures in Zoology by John Ashton - PDF Roomericdundee9No ratings yet
- VertebratesDocument63 pagesVertebratesgenusxyz100% (1)
- Diagnostic Test All SubjectsDocument23 pagesDiagnostic Test All SubjectsEMMANUEL CRUZNo ratings yet
- Vertebrates (Kingdom Animalia)Document130 pagesVertebrates (Kingdom Animalia)Nurl AinaNo ratings yet
- Vision IAS Prelims 2020 Test 18 SDocument34 pagesVision IAS Prelims 2020 Test 18 StriloksinghmeenaNo ratings yet
- 4 Entrance Model Questions 2078Document11 pages4 Entrance Model Questions 2078wits.academy89No ratings yet
- MammalsDocument1 pageMammalsHelena OmeNo ratings yet
- Zoo102 - Mammalia MetatheriaDocument37 pagesZoo102 - Mammalia MetatheriaSushmita Zen BanaagNo ratings yet
- Endocrines and Osmoregulation - A Comparative Account in Vertebrates (PDFDrive)Document300 pagesEndocrines and Osmoregulation - A Comparative Account in Vertebrates (PDFDrive)Hafez A MabroukNo ratings yet
- 3 Vertebrates ContDocument89 pages3 Vertebrates ConthakimNo ratings yet
- AL 3.2 - Selecting and Constructing Test Items and TasksDocument8 pagesAL 3.2 - Selecting and Constructing Test Items and TasksJessa ParedesNo ratings yet
- Kingdom Animalia Phylum Chordata-2Document68 pagesKingdom Animalia Phylum Chordata-2api-285078865No ratings yet
- MammalsDocument30 pagesMammalsmikinandaputraNo ratings yet
- The Class MammaliaDocument5 pagesThe Class MammaliaMohammad Raza RizviNo ratings yet
- Class Mammalia NotesDocument17 pagesClass Mammalia NotesSamuel TosinNo ratings yet
- Selecting and Constructing Test Items and Tasks - Acog, C - AnswersDocument8 pagesSelecting and Constructing Test Items and Tasks - Acog, C - AnswersCherilyn AcogNo ratings yet















![Evolution: What Darwin Did Not Know by Then..! [And the Origin of Species Through Species-Branding]](https://imgv2-1-f.scribdassets.com/img/word_document/270231362/149x198/aea4885cd2/1677109978?v=1)





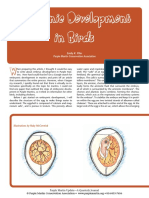


































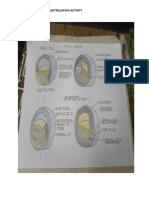






























Download as pdf or txt
You might also like
- Comparative Vertebrate AnatomyDocument11 pagesComparative Vertebrate AnatomyKean GB60% (5)
- Kiss NotesDocument38 pagesKiss NotesNalini Prakash100% (2)
- Textbook of Wild and Zoo AnimalsDocument247 pagesTextbook of Wild and Zoo AnimalsToha Putra100% (1)
- Chapter 1 - History of Zebrafish Resea - 2020 - The Zebrafish in Biomedical Rese PDFDocument12 pagesChapter 1 - History of Zebrafish Resea - 2020 - The Zebrafish in Biomedical Rese PDFNicolas BaronNo ratings yet
- 2012cooper Et Al Wires Development of Bat FlightDocument7 pages2012cooper Et Al Wires Development of Bat Flightapi-237346249No ratings yet
- Developmental Dynamics - 2013 - Sheng - Day 1 Chick DevelopmentDocument11 pagesDevelopmental Dynamics - 2013 - Sheng - Day 1 Chick DevelopmentYasir Iqbal KhanNo ratings yet
- Hendler 1975Document25 pagesHendler 1975byhz7wwdm4No ratings yet
- Integumentary SystemDocument51 pagesIntegumentary Systemmicklemagdy50No ratings yet
- Crustacean Larvae - Academic ScriptDocument12 pagesCrustacean Larvae - Academic ScriptAfridi ShaikhNo ratings yet
- Crustaceae Larval Forms PDFDocument12 pagesCrustaceae Larval Forms PDFAfridi ShaikhNo ratings yet
- Dvdy24087 5 Final As PublishedDocument12 pagesDvdy24087 5 Final As PublishedTahseen FatimaNo ratings yet
- Museum of Comparative Zoology, Harvard University, Cambridge, Massachusetts 02138Document12 pagesMuseum of Comparative Zoology, Harvard University, Cambridge, Massachusetts 02138Julio César Chávez GalarzaNo ratings yet
- Lesson 5 Birth Growth and Reproduction V5Document8 pagesLesson 5 Birth Growth and Reproduction V5JuanNo ratings yet
- Lab Act 1Document4 pagesLab Act 1Ostan Balanquit TeresaNo ratings yet
- Perspectives On The Use of The Baboon in Embryology and Teratology ResearchDocument18 pagesPerspectives On The Use of The Baboon in Embryology and Teratology ResearchYusraNo ratings yet
- Evolution: What Darwin Did Not Know by Then..! [And the Origin of Species Through Species-Branding]From EverandEvolution: What Darwin Did Not Know by Then..! [And the Origin of Species Through Species-Branding]No ratings yet
- 3rd Laboratory Activity AnuraDocument7 pages3rd Laboratory Activity AnuraZia Ammarah SaripNo ratings yet
- Elife 43828Document19 pagesElife 43828jaypasco79No ratings yet
- Zelfstudie Van Blastula Naar 3 KiemlagenDocument18 pagesZelfstudie Van Blastula Naar 3 KiemlagenCheyenne ProvoostNo ratings yet
- Medical Journal of Cell Biology Drosophila Melanogaster Research History, Breakthrough and PerspectivesDocument4 pagesMedical Journal of Cell Biology Drosophila Melanogaster Research History, Breakthrough and PerspectivesIgor DemićNo ratings yet
- Carril Et Al-2015-The Anatomical RecordDocument12 pagesCarril Et Al-2015-The Anatomical RecordrefestufuNo ratings yet
- Burung Matin PDFDocument2 pagesBurung Matin PDFRiskian Alfaro BabtistaNo ratings yet
- Altig & McDiarmid (2007) Morphological Diversity and Evolution of Egg and Clutch Structure in AmphibiansDocument33 pagesAltig & McDiarmid (2007) Morphological Diversity and Evolution of Egg and Clutch Structure in AmphibiansJuan Carlos SánchezNo ratings yet
- Oogenesis and Ovarian Histology of The South American Lizard Salvator MerianaeDocument14 pagesOogenesis and Ovarian Histology of The South American Lizard Salvator MerianaeValeria Garcia ValdezNo ratings yet
- Worksheet Q3 Week 5Document2 pagesWorksheet Q3 Week 5Jaybie TejadaNo ratings yet
- Prototheria: ClassificationDocument5 pagesPrototheria: ClassificationAalamgeer AnsariNo ratings yet
- Anatomy and Development of Oral Cavity and PharynxDocument14 pagesAnatomy and Development of Oral Cavity and PharynxGabriela MacoveanuNo ratings yet
- Evidence of Evolution - Module 6Document35 pagesEvidence of Evolution - Module 6Reign PabloNo ratings yet
- Matthews 1998Document9 pagesMatthews 1998osteopedcursoNo ratings yet
- The History of Vaginal BirthDocument11 pagesThe History of Vaginal BirthdardonnnNo ratings yet
- Evidence of EvolutionDocument15 pagesEvidence of Evolutionsurajkeshari9826No ratings yet
- Evolution of The Mammalian Neck From Developmental, Morpho-Functional, and Paleontological PerspectivesDocument11 pagesEvolution of The Mammalian Neck From Developmental, Morpho-Functional, and Paleontological PerspectivesKo KaNo ratings yet
- Embryologydevelopmental AnatomyDocument62 pagesEmbryologydevelopmental AnatomygirishNo ratings yet
- The Origin of The Turtle Body Plan EvideDocument19 pagesThe Origin of The Turtle Body Plan EvideXweuis Hekuos KweNo ratings yet
- Comparative Anatomy, Embryology & Vestigial StructuresDocument11 pagesComparative Anatomy, Embryology & Vestigial StructuresDaniel BerryNo ratings yet
- Basic Body Plan of Young Mammalian EmbryosDocument57 pagesBasic Body Plan of Young Mammalian EmbryosWin DierNo ratings yet
- Olfaction, Navigation, and The Origin of Isocortex: Francisco Aboitiz and Juan F. MontielDocument12 pagesOlfaction, Navigation, and The Origin of Isocortex: Francisco Aboitiz and Juan F. MontielmioclaenusNo ratings yet
- Blueprint of LifeDocument42 pagesBlueprint of Lifesophiehee1234No ratings yet
- Evidence of Evolution FinalDocument82 pagesEvidence of Evolution FinalCheskaNo ratings yet
- The Phylum CtenophoraDocument3 pagesThe Phylum CtenophoraTI Journals PublishingNo ratings yet
- Development of The Human Tail Bud and Splanchnic Mesenchyme: Original ArticleDocument7 pagesDevelopment of The Human Tail Bud and Splanchnic Mesenchyme: Original Articleproject-247758No ratings yet
- ZootaxaDocument17 pagesZootaxaoliviamuliawatideviNo ratings yet
- Group 4 Teacher'sDocument4 pagesGroup 4 Teacher'sAdrian GabrielNo ratings yet
- Regional Specification in The Early Embryo of The Brittle Star Ophiopholis AculeataDocument16 pagesRegional Specification in The Early Embryo of The Brittle Star Ophiopholis AculeataJoElunico QuezadaNo ratings yet
- Comisuritis Posterior 2Document8 pagesComisuritis Posterior 2VORTIZNo ratings yet
- Gastrulation in The Mouse Embryo: Ultrastructural and Molecular Aspects of Germ Layer MorphogenesisDocument28 pagesGastrulation in The Mouse Embryo: Ultrastructural and Molecular Aspects of Germ Layer MorphogenesisDIEGO FERNANDO URIBE JARAMILLONo ratings yet
- Order Myctophiformes: Blackchins and Lanternfishes by H. Geoffrey Moser and William WatsonDocument49 pagesOrder Myctophiformes: Blackchins and Lanternfishes by H. Geoffrey Moser and William WatsonAcuario Hatchery Los VilosNo ratings yet
- j.1463 6395.2008.00349.x20170103 12513 B3dicm With Cover Page v2Document12 pagesj.1463 6395.2008.00349.x20170103 12513 B3dicm With Cover Page v2Zulema EmirNo ratings yet
- Hendler 1986Document8 pagesHendler 1986byhz7wwdm4No ratings yet
- IESO Preparation Material (Paleontology)Document25 pagesIESO Preparation Material (Paleontology)Science Olympiad Blog67% (3)
- Larva of CrustaceanDocument9 pagesLarva of Crustaceanblloveraastha111No ratings yet
- Larva: Evolutionary TheoryDocument2 pagesLarva: Evolutionary TheoryEdron Bullo JrNo ratings yet
- Lecture 10-Coelomate BauplanDocument59 pagesLecture 10-Coelomate BauplanKago NgomaNo ratings yet
- Seijjjj 378777y36262626363. 4444 22373737374 A - U8ujb Vjhdjdjd. Bbwqwqwoo Rchins - As - Model - System - For - Studying - DevelopmentDocument7 pagesSeijjjj 378777y36262626363. 4444 22373737374 A - U8ujb Vjhdjdjd. Bbwqwqwoo Rchins - As - Model - System - For - Studying - DevelopmentRed DiggerNo ratings yet
- TEAM A BIo 180 Lab Learning Contract 1 Evidence of EvolutionDocument20 pagesTEAM A BIo 180 Lab Learning Contract 1 Evidence of EvolutionJames Carbonell Dela PeñaNo ratings yet
- Sed 202 Biological Science Research Tasks BDocument8 pagesSed 202 Biological Science Research Tasks BBryllie BautistaNo ratings yet
- Fullregenerationoftribasal Polypterusfin PNASDocument9 pagesFullregenerationoftribasal Polypterusfin PNASBagas Lantip PrakasaNo ratings yet
- GASTRULATION-ACTIVITY - DEV-BIO - Bio 102Document6 pagesGASTRULATION-ACTIVITY - DEV-BIO - Bio 102Quentin LopezNo ratings yet
- Donoghue Et Al-2014-PalaeontologyDocument16 pagesDonoghue Et Al-2014-PalaeontologyzhaNo ratings yet
- Introduction To The Zebrafish: MaterialsDocument9 pagesIntroduction To The Zebrafish: MaterialsDogNo ratings yet
- Pakistan Vertebrate FaunaDocument9 pagesPakistan Vertebrate FaunaNosheen MalikNo ratings yet
- Kingdom: Animals: Domain EukaryaDocument25 pagesKingdom: Animals: Domain EukaryaDoods GaldoNo ratings yet
- RTP Las English 5 Q3 Melc 2C - Summarizing Classification Text TypeDocument10 pagesRTP Las English 5 Q3 Melc 2C - Summarizing Classification Text TypePinaka ShungaNo ratings yet
- Kapocs Australia 5-6 PDFDocument21 pagesKapocs Australia 5-6 PDFMohamed HassanNo ratings yet
- Animal KingdomDocument67 pagesAnimal KingdomNaveen Krishna M.KNo ratings yet
- SDADASDASDASDDocument16 pagesSDADASDASDASDAngelaNo ratings yet
- A Manual of Mammalogy With Keys To Families of The World 3rd Edition Ebook PDFDocument62 pagesA Manual of Mammalogy With Keys To Families of The World 3rd Edition Ebook PDFandrea.davis76598% (46)
- 1 Intro To Exotics 7-2023Document56 pages1 Intro To Exotics 7-2023Hailey WilliamsNo ratings yet
- Origin of MammalsDocument26 pagesOrigin of Mammalsjai kumarNo ratings yet
- p.6 Paramount Integrated Science Notes Term 1Document152 pagesp.6 Paramount Integrated Science Notes Term 1Edward Biryetega100% (2)
- Interactive Textbook ScienceDocument464 pagesInteractive Textbook ScienceJose Manuel Rodriguez Cortes100% (3)
- 109Document14 pages109manhtuan15aNo ratings yet
- Britannica Illustrated Science Library - Mammals PDFDocument56 pagesBritannica Illustrated Science Library - Mammals PDFCupitu MadalinaNo ratings yet
- Curious Creatures in Zoology by John Ashton - PDF RoomDocument112 pagesCurious Creatures in Zoology by John Ashton - PDF Roomericdundee9No ratings yet
- VertebratesDocument63 pagesVertebratesgenusxyz100% (1)
- Diagnostic Test All SubjectsDocument23 pagesDiagnostic Test All SubjectsEMMANUEL CRUZNo ratings yet
- Vertebrates (Kingdom Animalia)Document130 pagesVertebrates (Kingdom Animalia)Nurl AinaNo ratings yet
- Vision IAS Prelims 2020 Test 18 SDocument34 pagesVision IAS Prelims 2020 Test 18 StriloksinghmeenaNo ratings yet
- 4 Entrance Model Questions 2078Document11 pages4 Entrance Model Questions 2078wits.academy89No ratings yet
- MammalsDocument1 pageMammalsHelena OmeNo ratings yet
- Zoo102 - Mammalia MetatheriaDocument37 pagesZoo102 - Mammalia MetatheriaSushmita Zen BanaagNo ratings yet
- Endocrines and Osmoregulation - A Comparative Account in Vertebrates (PDFDrive)Document300 pagesEndocrines and Osmoregulation - A Comparative Account in Vertebrates (PDFDrive)Hafez A MabroukNo ratings yet
- 3 Vertebrates ContDocument89 pages3 Vertebrates ConthakimNo ratings yet
- AL 3.2 - Selecting and Constructing Test Items and TasksDocument8 pagesAL 3.2 - Selecting and Constructing Test Items and TasksJessa ParedesNo ratings yet
- Kingdom Animalia Phylum Chordata-2Document68 pagesKingdom Animalia Phylum Chordata-2api-285078865No ratings yet
- MammalsDocument30 pagesMammalsmikinandaputraNo ratings yet
- The Class MammaliaDocument5 pagesThe Class MammaliaMohammad Raza RizviNo ratings yet
- Class Mammalia NotesDocument17 pagesClass Mammalia NotesSamuel TosinNo ratings yet
- Selecting and Constructing Test Items and Tasks - Acog, C - AnswersDocument8 pagesSelecting and Constructing Test Items and Tasks - Acog, C - AnswersCherilyn AcogNo ratings yet