
Hinton - 2014 - Histología y Genes Que Regulan El Crecimiento de La ATM
Hinton - 2014 - Histología y Genes Que Regulan El Crecimiento de La ATM
You might also like
- Eva M. Wojcik, Daniel F.I. Kurtycz, Dorothy L. Rosenthal - The Paris System For Reporting Urinary Cytology-Springer (2022)Document340 pagesEva M. Wojcik, Daniel F.I. Kurtycz, Dorothy L. Rosenthal - The Paris System For Reporting Urinary Cytology-Springer (2022)learta100% (2)
- NEO, April 01, 2021 Volume 22, Issue 4Document70 pagesNEO, April 01, 2021 Volume 22, Issue 4Majo EscobarNo ratings yet
- Orthopaedics and Trauma for Medical Students and Junior ResidentsFrom EverandOrthopaedics and Trauma for Medical Students and Junior ResidentsRating: 5 out of 5 stars5/5 (3)
- The Pediatric Mandible I. A Primer On Growth and DevelopmentDocument10 pagesThe Pediatric Mandible I. A Primer On Growth and DevelopmentJulián David Ríos ZuluagaNo ratings yet
- Embriologia Humana y Biologia Del Desarrollo CarlsonDocument14 pagesEmbriologia Humana y Biologia Del Desarrollo CarlsonJulio AbarzuaNo ratings yet
- 2018 - Development and Physiology of The Temporomandibular JointDocument9 pages2018 - Development and Physiology of The Temporomandibular JointJULIA SANFURGO VILLARROELNo ratings yet
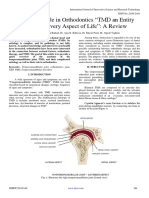
- TMD & Its Role in Orthodontics "TMD An Entity Affecting Every Aspect of Life" A ReviewDocument6 pagesTMD & Its Role in Orthodontics "TMD An Entity Affecting Every Aspect of Life" A ReviewInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- Bone DevelopmentDocument15 pagesBone DevelopmentVaibhav BhadageNo ratings yet
- Congenital Deformities and Developmental Abnormalities of The Mandibular Condyle in The Temporomandibular JointDocument8 pagesCongenital Deformities and Developmental Abnormalities of The Mandibular Condyle in The Temporomandibular JointMarcelaGutiérrezCarranzaNo ratings yet
- Temporomandibular Disorders. Part 1 PDFDocument11 pagesTemporomandibular Disorders. Part 1 PDFgunawanmulyana12No ratings yet
- OsteogensisDocument10 pagesOsteogensisDaniela OliverosNo ratings yet
- Paper 4Document9 pagesPaper 4Danna SalinasNo ratings yet
- Mouse GeneticDocument6 pagesMouse GeneticThanh Nhi NgôNo ratings yet
- Cranial Sutures: A Brief Review: SummaryDocument9 pagesCranial Sutures: A Brief Review: SummaryImane LoukdacheNo ratings yet
- 966ingenieria Celular CraneofacDocument14 pages966ingenieria Celular CraneofacFernandoFriasNo ratings yet
- Herb 2006Document7 pagesHerb 2006tatiana_gil_4No ratings yet
- Temporomandibular Joint: Aditi PGT 1 YearDocument27 pagesTemporomandibular Joint: Aditi PGT 1 Yearsachin mishraNo ratings yet
- Articular Disc Displacement Disorders and Its Prosthodontic Management: A Literature ReviewDocument9 pagesArticular Disc Displacement Disorders and Its Prosthodontic Management: A Literature ReviewTopan MuhamadNo ratings yet
- Osterix-Mcherry Transgenic Medaka For In: Vivo Imaging of Bone FormationDocument8 pagesOsterix-Mcherry Transgenic Medaka For In: Vivo Imaging of Bone Formationyui kirigayayuukiNo ratings yet
- Accepted Manuscript: Seminars in RoentgenologyDocument65 pagesAccepted Manuscript: Seminars in RoentgenologycooltomsNo ratings yet
- Why MechanobiologyDocument14 pagesWhy MechanobiologyNICOLAS CARVANo ratings yet
- TMJ Development PDFDocument10 pagesTMJ Development PDFVidya SenthuNo ratings yet
- 08 Badel PDFDocument10 pages08 Badel PDFHONGJYNo ratings yet
- Crecimiento y Desarrollo de La ATMDocument9 pagesCrecimiento y Desarrollo de La ATMMaría Fernanda Cea QuintanaNo ratings yet
- DeLaurier-2018-Wiley Interdisciplinary Reviews3A Developmental BiologyDocument37 pagesDeLaurier-2018-Wiley Interdisciplinary Reviews3A Developmental BiologyRaymundo Lopez NNo ratings yet
- A Single-Cell Transcriptional Atlas Reveals Resident Progenitor Cell Niche Functions in TMJ Disc Development and InjuryDocument18 pagesA Single-Cell Transcriptional Atlas Reveals Resident Progenitor Cell Niche Functions in TMJ Disc Development and InjuryThanh Nhi NgôNo ratings yet
- Evolution and Development of The Fish Jaw Skeleton: Wiley Interdisciplinary Reviews: Developmental Biology October 2018Document37 pagesEvolution and Development of The Fish Jaw Skeleton: Wiley Interdisciplinary Reviews: Developmental Biology October 2018Misskweenee DequitoNo ratings yet
- t3 Som 2014 2Document14 pagest3 Som 2014 2Julio AbarzuaNo ratings yet
- Basic Mechanisms in Craniofacial GrowthDocument9 pagesBasic Mechanisms in Craniofacial GrowthYady S. Londoño L.100% (1)
- Developmental Dynamics - 2008 - Hu - Effect of Bone Morphogenetic Protein Signaling On Development of The Jaw SkeletonDocument11 pagesDevelopmental Dynamics - 2008 - Hu - Effect of Bone Morphogenetic Protein Signaling On Development of The Jaw Skeletonhennysusanto18No ratings yet
- FisisDocument28 pagesFisisDaniel AguilarNo ratings yet
- Anatomy AtmDocument6 pagesAnatomy AtmNataly Mesa HernandezNo ratings yet
- V 014 Supp 02 A 015Document1 pageV 014 Supp 02 A 015Isaac Ooi Poh JitNo ratings yet
- World's Largest Science, Technology & Medicine Open Access Book PublisherDocument31 pagesWorld's Largest Science, Technology & Medicine Open Access Book PublisherAde Dian KaruniaNo ratings yet
- Buchtova 2007 Zoology II PDFDocument21 pagesBuchtova 2007 Zoology II PDFEduardo Panadero CuarteroNo ratings yet
- The Differential Roles of Periosteal and Capsular Functional Matrices in Orofacial GrowthDocument6 pagesThe Differential Roles of Periosteal and Capsular Functional Matrices in Orofacial GrowthCony Del CidNo ratings yet
- Keith1982 PDFDocument8 pagesKeith1982 PDFLakshmi Sarvani ValluriNo ratings yet
- Growth and Development 2Document10 pagesGrowth and Development 2Ali MONo ratings yet
- Peran Hormon Thyroid Pada CraniofacialDocument18 pagesPeran Hormon Thyroid Pada CraniofacialAnde Mangkuluhur Azhari ThalibbanNo ratings yet
- Biology of Bone Formation, Fracture Healing, and Distraction OsteogenesisDocument10 pagesBiology of Bone Formation, Fracture Healing, and Distraction OsteogenesisenviNo ratings yet
- Fcell 09 664168Document13 pagesFcell 09 664168Alya RahmadaniNo ratings yet
- Bedoya 2014Document10 pagesBedoya 2014Ortodoncia UnicocNo ratings yet
- 1525973984Document8 pages1525973984jrardcNo ratings yet
- 2-Referencias y Tecnicas Usadas en Clasificacion Hueso Alveolar - SeriwatanachaiDocument5 pages2-Referencias y Tecnicas Usadas en Clasificacion Hueso Alveolar - SeriwatanachaimnhjrtbnzbNo ratings yet
- The Epiphyseal Plate: Physiology, Anatomy, and TraumaDocument12 pagesThe Epiphyseal Plate: Physiology, Anatomy, and TraumaLaura Mérida CáceresNo ratings yet
- Articulo 1 CX DecimoDocument6 pagesArticulo 1 CX Decimodentist booksNo ratings yet
- Um Caminho para o Osso Moléculas Sinalizadoras e Fatores de TranscriçãoDocument15 pagesUm Caminho para o Osso Moléculas Sinalizadoras e Fatores de Transcriçãocursotplo2021No ratings yet
- The Differential Roles of Periosteal and Capsular Functional Matrices in Orofacial GrowthDocument6 pagesThe Differential Roles of Periosteal and Capsular Functional Matrices in Orofacial GrowthAaron Neal DmdNo ratings yet
- 11 Interdisciplinary Management of Dentofacial Deformity in Juvenile Idiopathic ArthritisDocument18 pages11 Interdisciplinary Management of Dentofacial Deformity in Juvenile Idiopathic ArthritiscdentistacesarmNo ratings yet
- Evolution, Epidemiology and Etiology of Temporomandibular Joint DisordersDocument6 pagesEvolution, Epidemiology and Etiology of Temporomandibular Joint DisordersakNo ratings yet
- Evolution, Epidemiology and Etiology of Temporomandibular Joint DisordersDocument6 pagesEvolution, Epidemiology and Etiology of Temporomandibular Joint DisordersakNo ratings yet
- لبنى زهيرDocument54 pagesلبنى زهيرdrshariquennazirNo ratings yet
- 2016 - Observing The Development of The ATM in EmbryonicDocument9 pages2016 - Observing The Development of The ATM in EmbryonicJULIA SANFURGO VILLARROELNo ratings yet
- The Human Meniscus: A Review of Anatomy, Function, Injury, and Advances in TreatmentDocument19 pagesThe Human Meniscus: A Review of Anatomy, Function, Injury, and Advances in TreatmentRazlivenaTintaNo ratings yet
- Macaca: Comparative Study of Cranial Growth in andDocument11 pagesMacaca: Comparative Study of Cranial Growth in andArbnor KicaNo ratings yet
- Tema 1 - Pulpa y Tejidos PerirradicularesDocument21 pagesTema 1 - Pulpa y Tejidos PerirradicularesLuz Maria Lopez PerezNo ratings yet
- TMJ in Embryonic and MiceDocument9 pagesTMJ in Embryonic and MiceThanh Nhi NgôNo ratings yet
- Growth and Development: Hereditary and Mechanical ModulationsDocument12 pagesGrowth and Development: Hereditary and Mechanical ModulationsPROSIG GERENTENo ratings yet
- Periodontal Ligament and Alveolar Bone in Health and Adaptation: Tooth MovementDocument8 pagesPeriodontal Ligament and Alveolar Bone in Health and Adaptation: Tooth MovementPentiya Vita AyuniNo ratings yet
- Cunningham Et Al 2016 Chapter 3Document17 pagesCunningham Et Al 2016 Chapter 3Rodrigo Vivero MirandaNo ratings yet
- TMJDocument66 pagesTMJmounika50% (2)
- Sporniak Tutak2011Document6 pagesSporniak Tutak2011Tarsardo marbunNo ratings yet
- Brochet - 2023 - Fotografía en Cirugía MaxilofacialDocument8 pagesBrochet - 2023 - Fotografía en Cirugía MaxilofacialMICHAEL ERNESTO QUINCHE RODRIGUEZNo ratings yet
- Badel 2007Document10 pagesBadel 2007MICHAEL ERNESTO QUINCHE RODRIGUEZNo ratings yet
- Golez - 2023 - Ortodoncia y Sensibilidad Dental - Revisión y Meta AnálisisDocument14 pagesGolez - 2023 - Ortodoncia y Sensibilidad Dental - Revisión y Meta AnálisisMICHAEL ERNESTO QUINCHE RODRIGUEZNo ratings yet
- Dallel 2019impact of Orthodontic Forces On Pulp Reaction A Systematic ReviewDocument6 pagesDallel 2019impact of Orthodontic Forces On Pulp Reaction A Systematic ReviewMICHAEL ERNESTO QUINCHE RODRIGUEZNo ratings yet
- Nosology and Classification of Genetic Skeletal Disorders - 2015 RevisionDocument24 pagesNosology and Classification of Genetic Skeletal Disorders - 2015 RevisionMariaYoanitaAstrianiNo ratings yet
- AchondroplasiaDocument9 pagesAchondroplasia9638391No ratings yet
- Citation IndexDocument5 pagesCitation IndexJoe WilsonNo ratings yet
- Nosology of Genetic Skeletal Disorders 2023 Revision - CompressedDocument46 pagesNosology of Genetic Skeletal Disorders 2023 Revision - CompressedAntoine NinoNo ratings yet
- Achondroplasia-CaseStudy FinalDocument18 pagesAchondroplasia-CaseStudy FinalJhovelNo ratings yet
- Achondroplasia DwarfismDocument12 pagesAchondroplasia Dwarfismandi firdha restuwatiNo ratings yet
- MSK Congenital DisordersDocument49 pagesMSK Congenital DisordersFarah Balqis BaragbahNo ratings yet
- Achondroplasia: A Comprehensive Clinical ReviewDocument49 pagesAchondroplasia: A Comprehensive Clinical ReviewdeviraNo ratings yet
- Achondroplasia - A Comprehensive Clinical Review 1-15Document15 pagesAchondroplasia - A Comprehensive Clinical Review 1-15jossua carlosamaNo ratings yet
- Tarsal CoalitionDocument7 pagesTarsal CoalitionFran coNo ratings yet
- Biology of Bone Formation, Fracture Healing, and Distraction OsteogenesisDocument10 pagesBiology of Bone Formation, Fracture Healing, and Distraction OsteogenesisenviNo ratings yet
- An Induced Pluripotent Stem Cell Line GZHMCi004 A Derived Fro - 2021 - Stem CeDocument4 pagesAn Induced Pluripotent Stem Cell Line GZHMCi004 A Derived Fro - 2021 - Stem CeFrankenstein MelancholyNo ratings yet
- Thanatoporic Dysplasia FIXDocument7 pagesThanatoporic Dysplasia FIXYudha GanesaNo ratings yet
- Grp.3 Bio Case Study of DwarfismDocument9 pagesGrp.3 Bio Case Study of DwarfismJhovelNo ratings yet
- Jurnal Sitologi UrineDocument10 pagesJurnal Sitologi UrineAjip JenNo ratings yet













































































Download as pdf or txt
You might also like
- Eva M. Wojcik, Daniel F.I. Kurtycz, Dorothy L. Rosenthal - The Paris System For Reporting Urinary Cytology-Springer (2022)Document340 pagesEva M. Wojcik, Daniel F.I. Kurtycz, Dorothy L. Rosenthal - The Paris System For Reporting Urinary Cytology-Springer (2022)learta100% (2)
- NEO, April 01, 2021 Volume 22, Issue 4Document70 pagesNEO, April 01, 2021 Volume 22, Issue 4Majo EscobarNo ratings yet
- Orthopaedics and Trauma for Medical Students and Junior ResidentsFrom EverandOrthopaedics and Trauma for Medical Students and Junior ResidentsRating: 5 out of 5 stars5/5 (3)
- The Pediatric Mandible I. A Primer On Growth and DevelopmentDocument10 pagesThe Pediatric Mandible I. A Primer On Growth and DevelopmentJulián David Ríos ZuluagaNo ratings yet
- Embriologia Humana y Biologia Del Desarrollo CarlsonDocument14 pagesEmbriologia Humana y Biologia Del Desarrollo CarlsonJulio AbarzuaNo ratings yet
- 2018 - Development and Physiology of The Temporomandibular JointDocument9 pages2018 - Development and Physiology of The Temporomandibular JointJULIA SANFURGO VILLARROELNo ratings yet
- TMD & Its Role in Orthodontics "TMD An Entity Affecting Every Aspect of Life" A ReviewDocument6 pagesTMD & Its Role in Orthodontics "TMD An Entity Affecting Every Aspect of Life" A ReviewInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- Bone DevelopmentDocument15 pagesBone DevelopmentVaibhav BhadageNo ratings yet
- Congenital Deformities and Developmental Abnormalities of The Mandibular Condyle in The Temporomandibular JointDocument8 pagesCongenital Deformities and Developmental Abnormalities of The Mandibular Condyle in The Temporomandibular JointMarcelaGutiérrezCarranzaNo ratings yet
- Temporomandibular Disorders. Part 1 PDFDocument11 pagesTemporomandibular Disorders. Part 1 PDFgunawanmulyana12No ratings yet
- OsteogensisDocument10 pagesOsteogensisDaniela OliverosNo ratings yet
- Paper 4Document9 pagesPaper 4Danna SalinasNo ratings yet
- Mouse GeneticDocument6 pagesMouse GeneticThanh Nhi NgôNo ratings yet
- Cranial Sutures: A Brief Review: SummaryDocument9 pagesCranial Sutures: A Brief Review: SummaryImane LoukdacheNo ratings yet
- 966ingenieria Celular CraneofacDocument14 pages966ingenieria Celular CraneofacFernandoFriasNo ratings yet
- Herb 2006Document7 pagesHerb 2006tatiana_gil_4No ratings yet
- Temporomandibular Joint: Aditi PGT 1 YearDocument27 pagesTemporomandibular Joint: Aditi PGT 1 Yearsachin mishraNo ratings yet
- Articular Disc Displacement Disorders and Its Prosthodontic Management: A Literature ReviewDocument9 pagesArticular Disc Displacement Disorders and Its Prosthodontic Management: A Literature ReviewTopan MuhamadNo ratings yet
- Osterix-Mcherry Transgenic Medaka For In: Vivo Imaging of Bone FormationDocument8 pagesOsterix-Mcherry Transgenic Medaka For In: Vivo Imaging of Bone Formationyui kirigayayuukiNo ratings yet
- Accepted Manuscript: Seminars in RoentgenologyDocument65 pagesAccepted Manuscript: Seminars in RoentgenologycooltomsNo ratings yet
- Why MechanobiologyDocument14 pagesWhy MechanobiologyNICOLAS CARVANo ratings yet
- TMJ Development PDFDocument10 pagesTMJ Development PDFVidya SenthuNo ratings yet
- 08 Badel PDFDocument10 pages08 Badel PDFHONGJYNo ratings yet
- Crecimiento y Desarrollo de La ATMDocument9 pagesCrecimiento y Desarrollo de La ATMMaría Fernanda Cea QuintanaNo ratings yet
- DeLaurier-2018-Wiley Interdisciplinary Reviews3A Developmental BiologyDocument37 pagesDeLaurier-2018-Wiley Interdisciplinary Reviews3A Developmental BiologyRaymundo Lopez NNo ratings yet
- A Single-Cell Transcriptional Atlas Reveals Resident Progenitor Cell Niche Functions in TMJ Disc Development and InjuryDocument18 pagesA Single-Cell Transcriptional Atlas Reveals Resident Progenitor Cell Niche Functions in TMJ Disc Development and InjuryThanh Nhi NgôNo ratings yet
- Evolution and Development of The Fish Jaw Skeleton: Wiley Interdisciplinary Reviews: Developmental Biology October 2018Document37 pagesEvolution and Development of The Fish Jaw Skeleton: Wiley Interdisciplinary Reviews: Developmental Biology October 2018Misskweenee DequitoNo ratings yet
- t3 Som 2014 2Document14 pagest3 Som 2014 2Julio AbarzuaNo ratings yet
- Basic Mechanisms in Craniofacial GrowthDocument9 pagesBasic Mechanisms in Craniofacial GrowthYady S. Londoño L.100% (1)
- Developmental Dynamics - 2008 - Hu - Effect of Bone Morphogenetic Protein Signaling On Development of The Jaw SkeletonDocument11 pagesDevelopmental Dynamics - 2008 - Hu - Effect of Bone Morphogenetic Protein Signaling On Development of The Jaw Skeletonhennysusanto18No ratings yet
- FisisDocument28 pagesFisisDaniel AguilarNo ratings yet
- Anatomy AtmDocument6 pagesAnatomy AtmNataly Mesa HernandezNo ratings yet
- V 014 Supp 02 A 015Document1 pageV 014 Supp 02 A 015Isaac Ooi Poh JitNo ratings yet
- World's Largest Science, Technology & Medicine Open Access Book PublisherDocument31 pagesWorld's Largest Science, Technology & Medicine Open Access Book PublisherAde Dian KaruniaNo ratings yet
- Buchtova 2007 Zoology II PDFDocument21 pagesBuchtova 2007 Zoology II PDFEduardo Panadero CuarteroNo ratings yet
- The Differential Roles of Periosteal and Capsular Functional Matrices in Orofacial GrowthDocument6 pagesThe Differential Roles of Periosteal and Capsular Functional Matrices in Orofacial GrowthCony Del CidNo ratings yet
- Keith1982 PDFDocument8 pagesKeith1982 PDFLakshmi Sarvani ValluriNo ratings yet
- Growth and Development 2Document10 pagesGrowth and Development 2Ali MONo ratings yet
- Peran Hormon Thyroid Pada CraniofacialDocument18 pagesPeran Hormon Thyroid Pada CraniofacialAnde Mangkuluhur Azhari ThalibbanNo ratings yet
- Biology of Bone Formation, Fracture Healing, and Distraction OsteogenesisDocument10 pagesBiology of Bone Formation, Fracture Healing, and Distraction OsteogenesisenviNo ratings yet
- Fcell 09 664168Document13 pagesFcell 09 664168Alya RahmadaniNo ratings yet
- Bedoya 2014Document10 pagesBedoya 2014Ortodoncia UnicocNo ratings yet
- 1525973984Document8 pages1525973984jrardcNo ratings yet
- 2-Referencias y Tecnicas Usadas en Clasificacion Hueso Alveolar - SeriwatanachaiDocument5 pages2-Referencias y Tecnicas Usadas en Clasificacion Hueso Alveolar - SeriwatanachaimnhjrtbnzbNo ratings yet
- The Epiphyseal Plate: Physiology, Anatomy, and TraumaDocument12 pagesThe Epiphyseal Plate: Physiology, Anatomy, and TraumaLaura Mérida CáceresNo ratings yet
- Articulo 1 CX DecimoDocument6 pagesArticulo 1 CX Decimodentist booksNo ratings yet
- Um Caminho para o Osso Moléculas Sinalizadoras e Fatores de TranscriçãoDocument15 pagesUm Caminho para o Osso Moléculas Sinalizadoras e Fatores de Transcriçãocursotplo2021No ratings yet
- The Differential Roles of Periosteal and Capsular Functional Matrices in Orofacial GrowthDocument6 pagesThe Differential Roles of Periosteal and Capsular Functional Matrices in Orofacial GrowthAaron Neal DmdNo ratings yet
- 11 Interdisciplinary Management of Dentofacial Deformity in Juvenile Idiopathic ArthritisDocument18 pages11 Interdisciplinary Management of Dentofacial Deformity in Juvenile Idiopathic ArthritiscdentistacesarmNo ratings yet
- Evolution, Epidemiology and Etiology of Temporomandibular Joint DisordersDocument6 pagesEvolution, Epidemiology and Etiology of Temporomandibular Joint DisordersakNo ratings yet
- Evolution, Epidemiology and Etiology of Temporomandibular Joint DisordersDocument6 pagesEvolution, Epidemiology and Etiology of Temporomandibular Joint DisordersakNo ratings yet
- لبنى زهيرDocument54 pagesلبنى زهيرdrshariquennazirNo ratings yet
- 2016 - Observing The Development of The ATM in EmbryonicDocument9 pages2016 - Observing The Development of The ATM in EmbryonicJULIA SANFURGO VILLARROELNo ratings yet
- The Human Meniscus: A Review of Anatomy, Function, Injury, and Advances in TreatmentDocument19 pagesThe Human Meniscus: A Review of Anatomy, Function, Injury, and Advances in TreatmentRazlivenaTintaNo ratings yet
- Macaca: Comparative Study of Cranial Growth in andDocument11 pagesMacaca: Comparative Study of Cranial Growth in andArbnor KicaNo ratings yet
- Tema 1 - Pulpa y Tejidos PerirradicularesDocument21 pagesTema 1 - Pulpa y Tejidos PerirradicularesLuz Maria Lopez PerezNo ratings yet
- TMJ in Embryonic and MiceDocument9 pagesTMJ in Embryonic and MiceThanh Nhi NgôNo ratings yet
- Growth and Development: Hereditary and Mechanical ModulationsDocument12 pagesGrowth and Development: Hereditary and Mechanical ModulationsPROSIG GERENTENo ratings yet
- Periodontal Ligament and Alveolar Bone in Health and Adaptation: Tooth MovementDocument8 pagesPeriodontal Ligament and Alveolar Bone in Health and Adaptation: Tooth MovementPentiya Vita AyuniNo ratings yet
- Cunningham Et Al 2016 Chapter 3Document17 pagesCunningham Et Al 2016 Chapter 3Rodrigo Vivero MirandaNo ratings yet
- TMJDocument66 pagesTMJmounika50% (2)
- Sporniak Tutak2011Document6 pagesSporniak Tutak2011Tarsardo marbunNo ratings yet
- Brochet - 2023 - Fotografía en Cirugía MaxilofacialDocument8 pagesBrochet - 2023 - Fotografía en Cirugía MaxilofacialMICHAEL ERNESTO QUINCHE RODRIGUEZNo ratings yet
- Badel 2007Document10 pagesBadel 2007MICHAEL ERNESTO QUINCHE RODRIGUEZNo ratings yet
- Golez - 2023 - Ortodoncia y Sensibilidad Dental - Revisión y Meta AnálisisDocument14 pagesGolez - 2023 - Ortodoncia y Sensibilidad Dental - Revisión y Meta AnálisisMICHAEL ERNESTO QUINCHE RODRIGUEZNo ratings yet
- Dallel 2019impact of Orthodontic Forces On Pulp Reaction A Systematic ReviewDocument6 pagesDallel 2019impact of Orthodontic Forces On Pulp Reaction A Systematic ReviewMICHAEL ERNESTO QUINCHE RODRIGUEZNo ratings yet
- Nosology and Classification of Genetic Skeletal Disorders - 2015 RevisionDocument24 pagesNosology and Classification of Genetic Skeletal Disorders - 2015 RevisionMariaYoanitaAstrianiNo ratings yet
- AchondroplasiaDocument9 pagesAchondroplasia9638391No ratings yet
- Citation IndexDocument5 pagesCitation IndexJoe WilsonNo ratings yet
- Nosology of Genetic Skeletal Disorders 2023 Revision - CompressedDocument46 pagesNosology of Genetic Skeletal Disorders 2023 Revision - CompressedAntoine NinoNo ratings yet
- Achondroplasia-CaseStudy FinalDocument18 pagesAchondroplasia-CaseStudy FinalJhovelNo ratings yet
- Achondroplasia DwarfismDocument12 pagesAchondroplasia Dwarfismandi firdha restuwatiNo ratings yet
- MSK Congenital DisordersDocument49 pagesMSK Congenital DisordersFarah Balqis BaragbahNo ratings yet
- Achondroplasia: A Comprehensive Clinical ReviewDocument49 pagesAchondroplasia: A Comprehensive Clinical ReviewdeviraNo ratings yet
- Achondroplasia - A Comprehensive Clinical Review 1-15Document15 pagesAchondroplasia - A Comprehensive Clinical Review 1-15jossua carlosamaNo ratings yet
- Tarsal CoalitionDocument7 pagesTarsal CoalitionFran coNo ratings yet
- Biology of Bone Formation, Fracture Healing, and Distraction OsteogenesisDocument10 pagesBiology of Bone Formation, Fracture Healing, and Distraction OsteogenesisenviNo ratings yet
- An Induced Pluripotent Stem Cell Line GZHMCi004 A Derived Fro - 2021 - Stem CeDocument4 pagesAn Induced Pluripotent Stem Cell Line GZHMCi004 A Derived Fro - 2021 - Stem CeFrankenstein MelancholyNo ratings yet
- Thanatoporic Dysplasia FIXDocument7 pagesThanatoporic Dysplasia FIXYudha GanesaNo ratings yet
- Grp.3 Bio Case Study of DwarfismDocument9 pagesGrp.3 Bio Case Study of DwarfismJhovelNo ratings yet
- Jurnal Sitologi UrineDocument10 pagesJurnal Sitologi UrineAjip JenNo ratings yet