Download as pdf or txt
You might also like
- Journal of Ethnopharmacology: Parminder Nain, Vipin Saini, Sunil Sharma, Jaspreet NainDocument7 pagesJournal of Ethnopharmacology: Parminder Nain, Vipin Saini, Sunil Sharma, Jaspreet NainAngie MenaNo ratings yet
- Chiết xuất lá atisô, như chất ức chế AKR1B1, làm giảm mức sorbitol trong thấu kính mắt chuột trong điều kiện glucose cao ex vivoDocument7 pagesChiết xuất lá atisô, như chất ức chế AKR1B1, làm giảm mức sorbitol trong thấu kính mắt chuột trong điều kiện glucose cao ex vivoLại Ngọc MaiNo ratings yet
- nrp-12-503Document9 pagesnrp-12-503ronahaniifah11No ratings yet
- Preparation, Characterization, and Antioxidant Properties of GallicDocument6 pagesPreparation, Characterization, and Antioxidant Properties of GallicAnonymous xGc8MRRysNo ratings yet
- Kim 2008Document6 pagesKim 2008tai phamNo ratings yet
- 5 - Structural and Functional Characterization of Polyphenols Isolated From Acerola Malpighia Emarginata DC FruitDocument8 pages5 - Structural and Functional Characterization of Polyphenols Isolated From Acerola Malpighia Emarginata DC FruitTALITA TALITANo ratings yet
- $RHZ3KN3Document39 pages$RHZ3KN3va.tmcbogorNo ratings yet
- Relative Protective Activities of Quercetin, Quercetin 3 Glucoside, and Rutin in Alcohol Induced Liver InjuryDocument9 pagesRelative Protective Activities of Quercetin, Quercetin 3 Glucoside, and Rutin in Alcohol Induced Liver InjuryMeow Vendo PermutoNo ratings yet
- Effect of Antihipoglycemic Sechium Edule Jacq. Swartz. Etanol Extract On Histopathologic Changes in Hyperglycemic Mus Musculus LDocument8 pagesEffect of Antihipoglycemic Sechium Edule Jacq. Swartz. Etanol Extract On Histopathologic Changes in Hyperglycemic Mus Musculus LAnindita Putri WardaniNo ratings yet
- 2013 Article 1220Document11 pages2013 Article 1220MeLissa AymNo ratings yet
- Ginkgo Biloba Extract in Alzheimer's Disease: From Action Mechanisms To Medical PracticeDocument17 pagesGinkgo Biloba Extract in Alzheimer's Disease: From Action Mechanisms To Medical PracticeTher RayNo ratings yet
- Research Article: Antifatigue Potential of Loquat Leaf Extract in Physical Stress in C2C12 Myotubes and in Vivo ModelsDocument12 pagesResearch Article: Antifatigue Potential of Loquat Leaf Extract in Physical Stress in C2C12 Myotubes and in Vivo ModelsjljsjoostenNo ratings yet
- 12-3-19 Vid 1 PDFDocument12 pages12-3-19 Vid 1 PDFchanduNo ratings yet
- 12-3-19 Vid 1 PDFDocument12 pages12-3-19 Vid 1 PDFchanduNo ratings yet
- A PyreneDocument22 pagesA PyreneArdina HendrianiNo ratings yet
- 006 Sudirman2022 - Seahorse Protein Hydrolysate Ameliorates ProinflammatoryDocument11 pages006 Sudirman2022 - Seahorse Protein Hydrolysate Ameliorates ProinflammatorySabri SudirmanNo ratings yet
- 1 s2.0 S0147651318301908 MainDocument11 pages1 s2.0 S0147651318301908 MainAndrea RetanaNo ratings yet
- 1 OnlineDocument11 pages1 OnlineArif ElsanNo ratings yet
- Protective Effect of Potato Peel Extract Against Carbon Tetrachloride-Induced Liver Injury in RatsDocument17 pagesProtective Effect of Potato Peel Extract Against Carbon Tetrachloride-Induced Liver Injury in RatsAzmi SevenfoldismNo ratings yet
- 500 - 1000 Pages For Book of Abstracts - I. INTERNATIONAL CONGRESS On MEDICINAL and AROMATIC PLANTS: "NATURAL and HEALTHY LIFE"Document500 pages500 - 1000 Pages For Book of Abstracts - I. INTERNATIONAL CONGRESS On MEDICINAL and AROMATIC PLANTS: "NATURAL and HEALTHY LIFE"Hasan Ibrahim Kozan50% (2)
- A-Glucosidase Inhibitors From The Seeds of Syagrus RomanzoffianaDocument6 pagesA-Glucosidase Inhibitors From The Seeds of Syagrus RomanzoffianaAlperay GirişbayNo ratings yet
- Nephroprotective Effects of Zingiber Zerumbet Smith Ethyl Acetate Extract Against Paracetamol-Induced Nephrotoxicity and Oxidative Stress in RatsDocument10 pagesNephroprotective Effects of Zingiber Zerumbet Smith Ethyl Acetate Extract Against Paracetamol-Induced Nephrotoxicity and Oxidative Stress in RatsIkhrimahNo ratings yet
- Dietary Antioxidant SupplementsDocument6 pagesDietary Antioxidant SupplementsNiar MarhaliNo ratings yet
- Molecules 24 00408Document24 pagesMolecules 24 00408everton.monteiroNo ratings yet
- 1 s2.0 S0895398821001069 MainDocument4 pages1 s2.0 S0895398821001069 MainShierly ShafrillaNo ratings yet
- Himani Paper 1-AndrologiaDocument13 pagesHimani Paper 1-AndrologiaLinguumNo ratings yet
- Artikel JaheDocument44 pagesArtikel Jahebunga nurcahayNo ratings yet
- Geriatrics 07 00064 v2Document9 pagesGeriatrics 07 00064 v2dandiokasubantaraNo ratings yet
- Low-Density Lipoprotein (LDL) - Antioxidant Lignans From Myristica Fragrans SeedsDocument5 pagesLow-Density Lipoprotein (LDL) - Antioxidant Lignans From Myristica Fragrans SeedsRakasiwi GalihNo ratings yet
- 1 s2.0 S1756464622000457 MainDocument10 pages1 s2.0 S1756464622000457 MainMiyyada AichaouiNo ratings yet
- Anti Cancer Mechanism and Possibility ofDocument4 pagesAnti Cancer Mechanism and Possibility ofOlivia Pebri RahayuNo ratings yet
- TMP 88 B5Document5 pagesTMP 88 B5FrontiersNo ratings yet
- Mechanistic Role of Epigallocatechin 3 Gallate in Regulation of The Antioxidant Markers in Ethanol Induced Liver Damage in MiceDocument10 pagesMechanistic Role of Epigallocatechin 3 Gallate in Regulation of The Antioxidant Markers in Ethanol Induced Liver Damage in MiceAllan WanderaNo ratings yet
- 1 s2.0 S0021967323006519 MainDocument6 pages1 s2.0 S0021967323006519 MainGonzalo Tereucan InaipilNo ratings yet
- High-Protein Bar As A Meal Replacement in Elite Sports Nutrition - A Pilot StudyDocument12 pagesHigh-Protein Bar As A Meal Replacement in Elite Sports Nutrition - A Pilot StudyvasilevnikolaiNo ratings yet
- Foods 09 01033 v2Document15 pagesFoods 09 01033 v2nguyendongthucNo ratings yet
- Antioxidant and Antidiabetic Effects of Gallic and Protocatechuic Acids: A Structure - Function PerspectiveDocument7 pagesAntioxidant and Antidiabetic Effects of Gallic and Protocatechuic Acids: A Structure - Function PerspectiveMentari ZikriNo ratings yet
- 1 s2.0 S2590157522002668 MainDocument14 pages1 s2.0 S2590157522002668 Mainazita nezamiNo ratings yet
- Latar Belakang Kayu ManisDocument11 pagesLatar Belakang Kayu ManisMuhammad YarhamNo ratings yet
- 41 81 1 SMDocument9 pages41 81 1 SMoffice8187No ratings yet
- Campos Et Al., 2014 Food ChemistryDocument7 pagesCampos Et Al., 2014 Food ChemistryMarceloGonzálezNo ratings yet
- 1 s2.0 S0753332223015068 MainDocument12 pages1 s2.0 S0753332223015068 Mainjair barruetaNo ratings yet
- Chaga Mushroom Extract Inhibits Oxidative Dna Damage in Human Lymphocytes As Assessed by Comet AssayDocument4 pagesChaga Mushroom Extract Inhibits Oxidative Dna Damage in Human Lymphocytes As Assessed by Comet Assayapi-85918191No ratings yet
- Protective Effects of Lagerstroemia Speciosa On 3-Morpholinosydnonimine (SIN-1) - Induced Oxidative Stress in HIT-T15 Pancreatic CellsDocument6 pagesProtective Effects of Lagerstroemia Speciosa On 3-Morpholinosydnonimine (SIN-1) - Induced Oxidative Stress in HIT-T15 Pancreatic CellsCurieThamerNo ratings yet
- Shi2010 PDFDocument17 pagesShi2010 PDFEBE THTNo ratings yet
- Original Article Influence of Industrial Sterilisation, Freezing and Steam Cooking On Antioxidant Properties of Green Peas and String BeansDocument8 pagesOriginal Article Influence of Industrial Sterilisation, Freezing and Steam Cooking On Antioxidant Properties of Green Peas and String Beansmarine2006No ratings yet
- Efectul de Prelucrare C-Âldur-Â Pe Detectarea PCR A Modificate Genetic (MG), Soia + N Produsele Din CarneDocument2 pagesEfectul de Prelucrare C-Âldur-Â Pe Detectarea PCR A Modificate Genetic (MG), Soia + N Produsele Din CarneStefan AdrianNo ratings yet
- Comparing The Phenolic Profile of Pilocarpus Pennatifolius Lem. ByHPLC-DAD-ESIMSnwith Respect To Authentication and Enzymeinhibition PotentialDocument11 pagesComparing The Phenolic Profile of Pilocarpus Pennatifolius Lem. ByHPLC-DAD-ESIMSnwith Respect To Authentication and Enzymeinhibition Potentialvlad_văluNo ratings yet
- Accepted Manuscript: DeserticolaDocument21 pagesAccepted Manuscript: DeserticolaFUCKMENo ratings yet
- Astaxantina (2011) - Astaxanthin Is Responsible For Antiglycoxidative Properties of Microalga Chlorella ZofingiensisDocument7 pagesAstaxantina (2011) - Astaxanthin Is Responsible For Antiglycoxidative Properties of Microalga Chlorella ZofingiensisamensetNo ratings yet
- Ginkgo BilobaDocument11 pagesGinkgo BilobaAnamaria RizacNo ratings yet
- Enzyme 1Document9 pagesEnzyme 1huynhthivietngan2002No ratings yet
- Pengaruh Ekstrak Rumput Laut (Euchema Sp.) Terhadap Histologi Sel Β Pankreas Pada Rattus Novergicus Galur Wistar Dengan Diabetes Mellitus Yang Di Injeksi Aloksan Kurniati Maya Sari, WdDocument8 pagesPengaruh Ekstrak Rumput Laut (Euchema Sp.) Terhadap Histologi Sel Β Pankreas Pada Rattus Novergicus Galur Wistar Dengan Diabetes Mellitus Yang Di Injeksi Aloksan Kurniati Maya Sari, Wdsri selfiana novita sariNo ratings yet
- Scientific PaperDocument14 pagesScientific PaperAhmed Hamdy KhattabNo ratings yet
- Molecules 22 00439Document16 pagesMolecules 22 00439Anonymous gzXHCYNo ratings yet
- Mahe 01Document5 pagesMahe 01mahemezNo ratings yet
- Antioxidant and Cytotoxic Activities of Centella Asiatica (L) UrbDocument9 pagesAntioxidant and Cytotoxic Activities of Centella Asiatica (L) UrbUwais AlNo ratings yet
- 1 s2.0 S0141813021017694 MainDocument7 pages1 s2.0 S0141813021017694 MainnathaloaNo ratings yet
- Nutrition and AgingDocument5 pagesNutrition and Agingbagus aldiNo ratings yet
- Kti-Ade Widya Kalsum - Xii Ipa 2Document36 pagesKti-Ade Widya Kalsum - Xii Ipa 2Ade Widya KalsumNo ratings yet
- MB ASCP Study DocumenttDocument12 pagesMB ASCP Study DocumenttLeeNo ratings yet
- Viton FDADocument6 pagesViton FDAAlejandro ZagalNo ratings yet
- Us 5959168Document9 pagesUs 5959168RafliNo ratings yet
- PCI Catalog - CompressedDocument20 pagesPCI Catalog - CompressedWalid HaouasNo ratings yet
- Fixation and Tissue ProcessingDocument7 pagesFixation and Tissue ProcessingMustafa AL-AsadsyNo ratings yet
- Product Overview: DRIVING INNOVATION in Natural-Based Lubricants For A Sustainable TomorrowDocument6 pagesProduct Overview: DRIVING INNOVATION in Natural-Based Lubricants For A Sustainable TomorrowLim Chee SiangNo ratings yet
- OIV - Compêndio Dos Métodos de Análise de Vinhos e MostosDocument43 pagesOIV - Compêndio Dos Métodos de Análise de Vinhos e MostosjmmfbeijosNo ratings yet
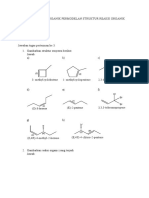
- 3-Methylcyclobutene 1 - Methylcyclopentene 2,3-Dimethyl-2-PenteneDocument3 pages3-Methylcyclobutene 1 - Methylcyclopentene 2,3-Dimethyl-2-PenteneFadilla AzhariNo ratings yet
- Ionic HW PDFDocument18 pagesIonic HW PDFKaustubh ThakerNo ratings yet
- Food Chemistry: I. Jerkovic, A. Kasum, Z. Marijanovic, C.I.G. TuberosoDocument10 pagesFood Chemistry: I. Jerkovic, A. Kasum, Z. Marijanovic, C.I.G. TuberosoKristina FilipovićNo ratings yet
- Assesment For Dna and RnaDocument1 pageAssesment For Dna and RnaJemebel NosaresNo ratings yet
- Advantages and Disadvantages of Techniques Used For WastewaterDocument11 pagesAdvantages and Disadvantages of Techniques Used For Wastewaterafham zulhusmiNo ratings yet
- CBLM - Produce Organic Concoction and ExtractsDocument43 pagesCBLM - Produce Organic Concoction and ExtractsRodel P. PilloNo ratings yet
- Lab Activity 5 Lipids: IUG, 2016 Dr. Tarek ZaidaDocument17 pagesLab Activity 5 Lipids: IUG, 2016 Dr. Tarek ZaidaJohanna Marie GantalaoNo ratings yet
- Synthetic Eyelash Extensions Are Made With Pbt. What Is PBT?Document3 pagesSynthetic Eyelash Extensions Are Made With Pbt. What Is PBT?kangge ngadownloadNo ratings yet
- Basic Concept of Organic Chemstry1 1Document40 pagesBasic Concept of Organic Chemstry1 1Raj ShresthaNo ratings yet
- Pdf&rendition 1Document9 pagesPdf&rendition 1ArchanaNo ratings yet
- Functions of BloodDocument8 pagesFunctions of BloodAndrés Lasso SNo ratings yet
- Preliminary Study On Thiamethoxam Degrading Bacteria Isolated From Corn PlantationDocument7 pagesPreliminary Study On Thiamethoxam Degrading Bacteria Isolated From Corn PlantationFazilah AriffinNo ratings yet
- Git Gastric JuiceDocument14 pagesGit Gastric JuiceSibatNo ratings yet
- Hardin S.Si, S.PD, M.PDDocument26 pagesHardin S.Si, S.PD, M.PDRadiatul Awalia AmirNo ratings yet
- Allen: Final Jee-Main Examination - June, 2022Document88 pagesAllen: Final Jee-Main Examination - June, 2022Ojas AlaiNo ratings yet
- Class 12th Cold Drinks Chemistry Investigatory Project Bhen Ke LaudeDocument23 pagesClass 12th Cold Drinks Chemistry Investigatory Project Bhen Ke LaudeMANJU KUMARINo ratings yet
- HisTrap HP 5x5mL Litdoc71502768 - 20150330002757Document28 pagesHisTrap HP 5x5mL Litdoc71502768 - 20150330002757Ana SmolkoNo ratings yet
- Lab ReportDocument3 pagesLab ReportJois SaturosNo ratings yet
- En 14331 PDFDocument5 pagesEn 14331 PDFWynona BasilioNo ratings yet
- Uop Unicracking Process For Hydrocracking: Donald AckelsonDocument12 pagesUop Unicracking Process For Hydrocracking: Donald AckelsonNguyễn Hoàng Bảo TrânNo ratings yet
- Auxiliary: Product ListDocument7 pagesAuxiliary: Product ListNguyễn Huy CườngNo ratings yet
- Organocadmium CompoundsDocument19 pagesOrganocadmium CompoundsGhazanfar Iqbal100% (2)