Download as pdf or txt
You might also like
- Test Bank For Campbell Essential Biology 7th by SimonDocument36 pagesTest Bank For Campbell Essential Biology 7th by Simonfuturityerlkingfd6d100% (33)
- Snorks Activity RevisedDocument8 pagesSnorks Activity RevisedFatima Viterbo Montalvo100% (1)
- Bioregulatory Systems Medicine Whitepaper PDFDocument75 pagesBioregulatory Systems Medicine Whitepaper PDFAlexander Diaz ZuletaNo ratings yet
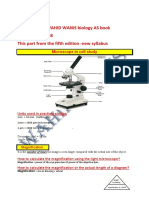
- Notes Dr. Wahid Wanis Biology ASFrom His BookDocument10 pagesNotes Dr. Wahid Wanis Biology ASFrom His BookBooks100% (1)
- Stewart 2016Document4 pagesStewart 2016alan.rangel.puenteNo ratings yet
- Beta3-Adrenoceptor Agonists Igawa 2010Document8 pagesBeta3-Adrenoceptor Agonists Igawa 2010Elysabet aristaNo ratings yet
- KellarTuanStockmeierAT 10012015Document11 pagesKellarTuanStockmeierAT 10012015Jean Pierre Chastre LuzaNo ratings yet
- Neuropeptide Y-Immunoreactive Nerve Terminals Parasympathetic Control of The Heart. IIIDocument10 pagesNeuropeptide Y-Immunoreactive Nerve Terminals Parasympathetic Control of The Heart. IIILuammmNo ratings yet
- 2020 The RAS in HypertensionDocument11 pages2020 The RAS in HypertensionPeter Albeiro Falla CortesNo ratings yet
- PTX 3Document9 pagesPTX 3M Ali AdrianNo ratings yet
- Clinical Handbook of Cardiac ElectrophysiologyFrom EverandClinical Handbook of Cardiac ElectrophysiologyBenedict M. GloverNo ratings yet
- B3 Adrenergic ReceptorDocument8 pagesB3 Adrenergic ReceptorlauraNo ratings yet
- ForebrainDocument26 pagesForebrainYixin TongNo ratings yet
- Pharmacology and Therapeutics of Omega-3 Polyunsaturated Fatty Acids in Chronic Inflammatory DiseaseDocument11 pagesPharmacology and Therapeutics of Omega-3 Polyunsaturated Fatty Acids in Chronic Inflammatory DiseasePeterJLYNo ratings yet
- BJP5 v. 170Document45 pagesBJP5 v. 170LourdesNo ratings yet
- Pulmonary Veno-Occlusive Disease and Pulmonary Hypertension in Dogs: Striking Similarities To The Human ConditionDocument4 pagesPulmonary Veno-Occlusive Disease and Pulmonary Hypertension in Dogs: Striking Similarities To The Human Conditionsebastian castro chavezNo ratings yet
- Clinical BiochemistryDocument5 pagesClinical BiochemistryRachmatu Bill MultazamNo ratings yet
- Interdependent Feedback Regulation of Breathing by The Carotid Bodies and The Retrotrapezoid NucleusDocument14 pagesInterdependent Feedback Regulation of Breathing by The Carotid Bodies and The Retrotrapezoid NucleusLezard DomiNo ratings yet
- A Dual Thrombin Receptor System For Platelet Activation: Nature September 1998Document6 pagesA Dual Thrombin Receptor System For Platelet Activation: Nature September 1998Kamonashis Sarkar TendulNo ratings yet
- Estudio Doppler de La Arteria Carótida Común Izquierda Con Estenosis PreoclusivaDocument5 pagesEstudio Doppler de La Arteria Carótida Común Izquierda Con Estenosis PreoclusivaJosè Julio Cejas SànchezNo ratings yet
- ASL Perfusion of The Brain Emerging Clinical ApplicationDocument20 pagesASL Perfusion of The Brain Emerging Clinical ApplicationpoppyNo ratings yet
- Reviews of Physiology, Biochemistry and Pharmacology 162Document126 pagesReviews of Physiology, Biochemistry and Pharmacology 162SolomonNo ratings yet
- Association of Serum PentraxinDocument11 pagesAssociation of Serum PentraxinWilliam KamarullahNo ratings yet
- B1-Adrenergic Receptors Increase UCP1 in Human MADS Brown Adipocytesand Rescue Cold-Acclimated B3-Adrenergic Receptor-Knockout Mice Vianonshivering ThermogenesisDocument11 pagesB1-Adrenergic Receptors Increase UCP1 in Human MADS Brown Adipocytesand Rescue Cold-Acclimated B3-Adrenergic Receptor-Knockout Mice Vianonshivering ThermogenesisAlfin M. AbdillahNo ratings yet
- Purinergic Receptors in The Carotid Body As A New Drug Target For Controlling HypertensionDocument12 pagesPurinergic Receptors in The Carotid Body As A New Drug Target For Controlling HypertensiongpatwallahNo ratings yet
- Interaction of Nerve Agent Antidotes With Cholinergic SystemsDocument12 pagesInteraction of Nerve Agent Antidotes With Cholinergic Systemsefrahmaa watiNo ratings yet
- Grant's Atlas of Anatomy Grant's Dissector: JAMA The Journal of The American Medical Association January 2008Document6 pagesGrant's Atlas of Anatomy Grant's Dissector: JAMA The Journal of The American Medical Association January 2008valmondsikiNo ratings yet
- Fauconnier2007 Insulin TRPC3Document10 pagesFauconnier2007 Insulin TRPC3Giulia AndreeaNo ratings yet
- Release 2+ and Independently of Sarcoplasmic Reticulum Ca Mediated TRPC3 Channel Activation Receptor 3 Constricts Cerebral Arteries Via IP 3 IPDocument18 pagesRelease 2+ and Independently of Sarcoplasmic Reticulum Ca Mediated TRPC3 Channel Activation Receptor 3 Constricts Cerebral Arteries Via IP 3 IPicanadaaNo ratings yet
- Sabevetal PDFDocument5 pagesSabevetal PDFDimas MedelluNo ratings yet
- Distinct From That of The Broad SpectrumDocument12 pagesDistinct From That of The Broad SpectrumPatricia T. WanaraNo ratings yet
- CARDIAC HEMANGIOMA IN A CORN SNAKE (PANTHEROPHIS GUTTATUS) - Stumpel2012Document8 pagesCARDIAC HEMANGIOMA IN A CORN SNAKE (PANTHEROPHIS GUTTATUS) - Stumpel2012Bruno Do CarmoNo ratings yet
- Ijms 16 14997Document12 pagesIjms 16 14997antonietohNo ratings yet
- Site-Specific O-Glycosylation by Polypeptide Galnac-Transferase T2 Co-Regulates Beta1-Adrenergic Receptor N-Terminal CleavageDocument27 pagesSite-Specific O-Glycosylation by Polypeptide Galnac-Transferase T2 Co-Regulates Beta1-Adrenergic Receptor N-Terminal CleavageHamayun KhanNo ratings yet
- Biology of Prostate Specific Antigen PDFDocument9 pagesBiology of Prostate Specific Antigen PDFKaren BotiaNo ratings yet
- Vasopressin Increases Phosphorylation of Ser84 and Ser486 in Slc14a2 Collecting Duct Urea TransportersDocument10 pagesVasopressin Increases Phosphorylation of Ser84 and Ser486 in Slc14a2 Collecting Duct Urea TransportersAlmas TNo ratings yet
- Therapeutic Uses of Antiepileptic Drugs. in Non-Epileptic DisordersDocument9 pagesTherapeutic Uses of Antiepileptic Drugs. in Non-Epileptic DisordersFabiola NogaNo ratings yet
- J Vet Emergen Crit Care - 2012 - Brainard - RECOVER Evidence and Knowledge Gap Analysis On Veterinary CPR Part 5Document20 pagesJ Vet Emergen Crit Care - 2012 - Brainard - RECOVER Evidence and Knowledge Gap Analysis On Veterinary CPR Part 5sebastian Jimenez PNo ratings yet
- CovidDocument12 pagesCovidSamuel GasparNo ratings yet
- Sacubitril Valsartan The Role of NeprilyDocument11 pagesSacubitril Valsartan The Role of NeprilySalvanabilaNo ratings yet
- 1 s2.0 S0021915021003269 MainDocument1 page1 s2.0 S0021915021003269 MainIoanna NikdimaNo ratings yet
- European Journal of Preventive Cardiology-2016-Groehs-2047487316654025Document11 pagesEuropean Journal of Preventive Cardiology-2016-Groehs-2047487316654025Michelly MachadoNo ratings yet
- Stroke: Literature Synopses: Basic ScienceDocument3 pagesStroke: Literature Synopses: Basic ScienceFajar Rudy QimindraNo ratings yet
- Extended Atrial Conduction System Characterised by The Expression of The HCN4 Channel and Connexin45Document11 pagesExtended Atrial Conduction System Characterised by The Expression of The HCN4 Channel and Connexin45CARLOS FRANCISCO MANTILLA MONTALVONo ratings yet
- Echocardiographic Assessment of Cardiac Structure and Function in Great Apes: A Practical GuideDocument17 pagesEchocardiographic Assessment of Cardiac Structure and Function in Great Apes: A Practical GuideMichael GregoryNo ratings yet
- Hepatic Encephalopathy in Dogs and Cats: State of The Art ReviewDocument17 pagesHepatic Encephalopathy in Dogs and Cats: State of The Art ReviewGuadalupe Cristina Chuchón CáceresNo ratings yet
- Benefits of Use of Power Lab in Practica PDFDocument4 pagesBenefits of Use of Power Lab in Practica PDFRoy HarperNo ratings yet
- TMP 3 CA1Document7 pagesTMP 3 CA1FrontiersNo ratings yet
- 2014 - Morfometria Cardíaca em CoelhosDocument8 pages2014 - Morfometria Cardíaca em CoelhosMagnoDosSantosRozaNo ratings yet
- Sistema de Información Científica: Red de Revistas Científicas de América Latina, El Caribe, España y PortugalDocument7 pagesSistema de Información Científica: Red de Revistas Científicas de América Latina, El Caribe, España y PortugalJ Roberto Meza OntiverosNo ratings yet
- 朱家权氯沙坦猪模型Document10 pages朱家权氯沙坦猪模型shentujin1997No ratings yet
- Pugazhendhi Vijayaraman Cardiac Conduction SystemDocument30 pagesPugazhendhi Vijayaraman Cardiac Conduction SystemMariaVirginiaNo ratings yet
- Sensory Hepatic NervesDocument9 pagesSensory Hepatic Nervescc vereNo ratings yet
- Wang2023-AKR1C3 & ASTHMADocument9 pagesWang2023-AKR1C3 & ASTHMAFajri FebrianNo ratings yet
- Stereotaxic Gene Delivery in The Rodent BrainDocument10 pagesStereotaxic Gene Delivery in The Rodent BrainDong DongNo ratings yet
- April: Jurg TschoppDocument4 pagesApril: Jurg TschoppactivnetNo ratings yet
- Parathyroid Hormone Coming of Age in Clinical MedDocument5 pagesParathyroid Hormone Coming of Age in Clinical MedSaad uddinNo ratings yet
- Camargo 2018 (Pathology, Pathophysio, Diagnosis, Symptomps, Treatment)Document6 pagesCamargo 2018 (Pathology, Pathophysio, Diagnosis, Symptomps, Treatment)Rizki Muhammad RanandaNo ratings yet
- The Renin Angiotensin System in Cardiovascular DiseaseDocument453 pagesThe Renin Angiotensin System in Cardiovascular DiseaseaslikandilNo ratings yet
- 2021 - Article - New Therapies For Lowering Triglyceride-Rich LipoproteinsDocument14 pages2021 - Article - New Therapies For Lowering Triglyceride-Rich Lipoproteinsnguyentruclam002No ratings yet
- Expression of A Breast-Cancer-Associated Protein (Ps2) in Human Neuro-Endocrine TumoursDocument5 pagesExpression of A Breast-Cancer-Associated Protein (Ps2) in Human Neuro-Endocrine TumoursSarly FebrianaNo ratings yet
- Hepatobiliary Excretion of Berberine: Pi-Lo Tsai and Tung-Hu TsaiDocument8 pagesHepatobiliary Excretion of Berberine: Pi-Lo Tsai and Tung-Hu TsaizebchemistNo ratings yet
- Biochemical and Biophysical Research CommunicationsDocument6 pagesBiochemical and Biophysical Research CommunicationsMuhammad AzharNo ratings yet
- Posterior Circulation Stroke: Advances in Understanding and ManagementFrom EverandPosterior Circulation Stroke: Advances in Understanding and ManagementJong S. KimNo ratings yet
- NIH Public Access: Author ManuscriptDocument23 pagesNIH Public Access: Author ManuscriptAlexander Diaz ZuletaNo ratings yet
- Genoma 1Document12 pagesGenoma 1Alexander Diaz ZuletaNo ratings yet
- Genoma 1Document12 pagesGenoma 1Alexander Diaz ZuletaNo ratings yet
- Disast PDFDocument6 pagesDisast PDFAlexander Diaz ZuletaNo ratings yet
- Disclosures: Kori Dewing, DNP, ARNP Virginia Mason Medical Center October 29, 2013Document3 pagesDisclosures: Kori Dewing, DNP, ARNP Virginia Mason Medical Center October 29, 2013Alexander Diaz ZuletaNo ratings yet
- Binding Immunoglobulin During DiseaseDocument12 pagesBinding Immunoglobulin During DiseaseAGLNo ratings yet
- Asumbi Girls High School: Biology Form Four Revision Introduction To BiologyDocument13 pagesAsumbi Girls High School: Biology Form Four Revision Introduction To Biologykaranjamary228No ratings yet
- Microbiology SyllabusDocument2 pagesMicrobiology Syllabusapi-3704804No ratings yet
- JPH 12720Document9 pagesJPH 12720tasha_abollaNo ratings yet
- Chapter 5 Bio f4Document20 pagesChapter 5 Bio f4Madyha AzmiNo ratings yet
- Science, Technology and Society: An Overview: Module 1, BI 140Document23 pagesScience, Technology and Society: An Overview: Module 1, BI 140Roberto Velasco MabulacNo ratings yet
- Chapter 10 - Nutrition and Gas Exchange in Plants: Learning ObjectivesDocument14 pagesChapter 10 - Nutrition and Gas Exchange in Plants: Learning Objectivesthanks btNo ratings yet
- 1.1 Role of Organic Drug Molecules Unit 1Document16 pages1.1 Role of Organic Drug Molecules Unit 1Amarnath SahNo ratings yet
- (6P) Basic Concepts in Immunity and InflammationDocument6 pages(6P) Basic Concepts in Immunity and InflammationNegrus Stefan100% (1)
- HDP To Lose Weight For Obesity TeenagersDocument8 pagesHDP To Lose Weight For Obesity TeenagersAnis Syazwina SalmanNo ratings yet
- INNO-LiPA HBV Multi-DR PDFDocument9 pagesINNO-LiPA HBV Multi-DR PDFJorge SantosNo ratings yet
- Body & Society: Biology's Gift: Interrogating The Turn To AffectDocument29 pagesBody & Society: Biology's Gift: Interrogating The Turn To AffectKauan NegriNo ratings yet
- Brief History of Human Blood GroupsDocument6 pagesBrief History of Human Blood GroupsTalhaNo ratings yet
- Inflamation AssignmentDocument59 pagesInflamation AssignmentAnonymous GRUjCVedhNo ratings yet
- Wise, King - 2008 - Mechanisms of Tooth Eruption and Orthodontic Tooth Movement PDFDocument22 pagesWise, King - 2008 - Mechanisms of Tooth Eruption and Orthodontic Tooth Movement PDFNeiba RomeroNo ratings yet
- LEC LE 3 Part 2 Stoichiometry of Growth and Product FormationDocument27 pagesLEC LE 3 Part 2 Stoichiometry of Growth and Product FormationRaymond Fuentes100% (1)
- Bio PBL F5 - HerbariumDocument14 pagesBio PBL F5 - HerbariumYu Han Ong100% (1)
- 2022 2024 SyllabusDocument33 pages2022 2024 Syllabus15jirantanin TapNo ratings yet
- 1 3 3 Evidence ReportDocument11 pages1 3 3 Evidence Reportapi-296590894No ratings yet
- The Human Life Begins at ConceptionDocument12 pagesThe Human Life Begins at ConceptionDavid KrogerNo ratings yet
- SC 102 Course Outline 1st Sem 2018Document3 pagesSC 102 Course Outline 1st Sem 2018Jocelle Ann RicablancaNo ratings yet
- Establishing Horticultural ProjectDocument8 pagesEstablishing Horticultural ProjectWILMOR A. DIONESNo ratings yet
- Affinity Chromatography Group 2 2bsmt3Document20 pagesAffinity Chromatography Group 2 2bsmt3nicole rebanalNo ratings yet
- 16.1 Asexual ReproductionDocument22 pages16.1 Asexual ReproductionEFFERUL MIFZALNo ratings yet
- Ebola Virus DiseaseDocument46 pagesEbola Virus DiseaseFatymah Azzahra RirinNo ratings yet
- Plant KingdomDocument6 pagesPlant Kingdomrautshreyash22No ratings yet
- Urban Ecology Australia - Do We Fit On The PlanetDocument9 pagesUrban Ecology Australia - Do We Fit On The PlanetRique BenitesNo ratings yet