
A Neural Code For Time and Space in The Human Brain
A Neural Code For Time and Space in The Human Brain
You might also like
- Gao Et Al. - 2020 - Neuronal Timescales Are Functionally Dynamic and SDocument26 pagesGao Et Al. - 2020 - Neuronal Timescales Are Functionally Dynamic and Srodrigo carlos carrilloNo ratings yet
- Fernando - Eichenbaum2017Document12 pagesFernando - Eichenbaum2017fernandogfcnsNo ratings yet
- "Inner GPS", A Far-Reaching in Uence in Brain Research-For The Nobel Prize in Physiology or Medicine 2014Document3 pages"Inner GPS", A Far-Reaching in Uence in Brain Research-For The Nobel Prize in Physiology or Medicine 2014mihaela irofteNo ratings yet
- Complexity of Cortical Wave Patterns of The Wake Mouse CortexDocument13 pagesComplexity of Cortical Wave Patterns of The Wake Mouse CortexgrviteNo ratings yet
- Lusk, 2016. Cerebellar Hippocampal and Striatal Time CellsDocument7 pagesLusk, 2016. Cerebellar Hippocampal and Striatal Time CellsAnaMaríaMalagónLNo ratings yet
- A Multi-Modal Parcellation of Human Cerebral Cortex: Europe PMC Funders GroupDocument25 pagesA Multi-Modal Parcellation of Human Cerebral Cortex: Europe PMC Funders GroupYu ZhaoNo ratings yet
- Buzsaki 2004 Nat NeurosciDocument7 pagesBuzsaki 2004 Nat Neuroscihussin elrashidyNo ratings yet
- Tes 111Document2 pagesTes 111Gramy LatumetenNo ratings yet
- Large-Scale Neural Recordings Call For New Insights To Link Brain and BehaviorDocument9 pagesLarge-Scale Neural Recordings Call For New Insights To Link Brain and BehaviorMoshe AlonsoNo ratings yet
- On Time and Space in The Brain - A Relativistic Pseudo-Diffusion FrameworkDocument14 pagesOn Time and Space in The Brain - A Relativistic Pseudo-Diffusion FrameworkGeorge LazarNo ratings yet
- CorrelacionDocument5 pagesCorrelacionlinne25No ratings yet
- Human Premotor Areas Parse Sequences Into Their Spatial and Temporal FeaturesDocument23 pagesHuman Premotor Areas Parse Sequences Into Their Spatial and Temporal FeaturesmoiNo ratings yet
- Neuronal Representation of Saccadic Error in Macaque Posterior Parietal Cortex (PPC)Document17 pagesNeuronal Representation of Saccadic Error in Macaque Posterior Parietal Cortex (PPC)Zeus HeraklesNo ratings yet
- 2015 - Nielson Et Al. - Human Hippocampus Represents Space and Time During Retrieval of Real-World MemoriesDocument6 pages2015 - Nielson Et Al. - Human Hippocampus Represents Space and Time During Retrieval of Real-World MemoriesNityananda PortelladaNo ratings yet
- TMP 4 FB0Document11 pagesTMP 4 FB0FrontiersNo ratings yet
- The Elusive Concept of Brain Connectivity: Barry HorwitzDocument5 pagesThe Elusive Concept of Brain Connectivity: Barry HorwitzParisa SattarNo ratings yet
- Ciliberti Et Al., 2018Document14 pagesCiliberti Et Al., 2018carlos rosalesNo ratings yet
- György Buzsáki - The Brain From Inside Out-Oxford University Press, USA (2019)Document20 pagesGyörgy Buzsáki - The Brain From Inside Out-Oxford University Press, USA (2019)juannnnNo ratings yet
- Paz Gelbard-Sagiv Fried2010-Hippocampal Substrate For Linking Successive EventsDocument6 pagesPaz Gelbard-Sagiv Fried2010-Hippocampal Substrate For Linking Successive Eventsiulia andreeaNo ratings yet
- Theta-Phase Dependent Neuronal Coding During Sequence Learning in Human Single NeuronsDocument12 pagesTheta-Phase Dependent Neuronal Coding During Sequence Learning in Human Single NeuronsWalter HuNo ratings yet
- Journal Pcbi 1011801Document30 pagesJournal Pcbi 1011801shivang patelNo ratings yet
- An Evolving Perspective On The Dynamic Brain - Notes From The Brain Conference On Dynamics of The Brain - Temporal Aspects of ComputationDocument22 pagesAn Evolving Perspective On The Dynamic Brain - Notes From The Brain Conference On Dynamics of The Brain - Temporal Aspects of ComputationRajsuryan SinghNo ratings yet
- Communication From The Cerebellum To The Neocortex During Sleep SpindlesDocument1 pageCommunication From The Cerebellum To The Neocortex During Sleep SpindlesDavid NazarovNo ratings yet
- Temporal EstimationDocument25 pagesTemporal EstimationangieNo ratings yet
- Sereno Et Al., 2020Document6 pagesSereno Et Al., 2020elinNo ratings yet
- Large and Fast Human Pyramidal Neurons Associate With IntelligenceDocument21 pagesLarge and Fast Human Pyramidal Neurons Associate With Intelligenceauf ayeNo ratings yet
- A Cortical Neural Prosthesis For Restoring and Enhancing Memory - Theodore W Berger - Dong Song - Vasilis Z Marmarelis - Et Al - JNeuralEng - 2011 - 1741-2552 - 8!4!046017Document12 pagesA Cortical Neural Prosthesis For Restoring and Enhancing Memory - Theodore W Berger - Dong Song - Vasilis Z Marmarelis - Et Al - JNeuralEng - 2011 - 1741-2552 - 8!4!046017Satyam NegiNo ratings yet
- 174 The Brain From Inside Out 135: Chapter 5. Internalization of ExperienceDocument20 pages174 The Brain From Inside Out 135: Chapter 5. Internalization of ExperiencejuannnnNo ratings yet
- Integrated Neurophotonics Toward Dense Volumetric Interrogation of Brain Circuit Activity-At Depth and in Real Time - 1-s2.0-S0896627320307674-MainDocument2 pagesIntegrated Neurophotonics Toward Dense Volumetric Interrogation of Brain Circuit Activity-At Depth and in Real Time - 1-s2.0-S0896627320307674-MainGeneration GenerationNo ratings yet
- Cognitive Map NobelDocument2 pagesCognitive Map NobelSakditad SaowapaNo ratings yet
- Conectoma Mosca Prepublicación PDFDocument76 pagesConectoma Mosca Prepublicación PDFSofia TombaNo ratings yet
- Using Goal-Driven Deep Learning Models To Understand The Sensory CortexDocument10 pagesUsing Goal-Driven Deep Learning Models To Understand The Sensory CortexRajsuryan SinghNo ratings yet
- Tarantino Et Al 2010Document10 pagesTarantino Et Al 2010jsaccuzzoNo ratings yet
- Review: Conscious Processing and The Global Neuronal Workspace HypothesisDocument23 pagesReview: Conscious Processing and The Global Neuronal Workspace HypothesisSebastian Gonzalez MazoNo ratings yet
- Conscious Visual PerceptionDocument46 pagesConscious Visual PerceptionDa CuNo ratings yet
- Large-Scale Recording of Neuronal Ensembles: PerspectiveDocument6 pagesLarge-Scale Recording of Neuronal Ensembles: PerspectiveMatheus PortelaNo ratings yet
- Sperry Roger Ecs 2003Document7 pagesSperry Roger Ecs 2003Johnmark ObligarNo ratings yet
- Millisecond by Millisecond, Year by Year: Normative EEG Microstates and Developmental StagesDocument5 pagesMillisecond by Millisecond, Year by Year: Normative EEG Microstates and Developmental Stagesscribd.com@jperla.comNo ratings yet
- 1 s2.0 S0092867420310771 MainDocument38 pages1 s2.0 S0092867420310771 MainmyczhNo ratings yet
- Silences Spikes and Bursts Three-Part Knot of TheDocument15 pagesSilences Spikes and Bursts Three-Part Knot of TheUlises ChialvaNo ratings yet
- Mapping Neuronal DiversityDocument2 pagesMapping Neuronal DiversityJohn MistryNo ratings yet
- Simplicial and Topological Descriptions of Human Brain DynamicsDocument20 pagesSimplicial and Topological Descriptions of Human Brain DynamicsJosue VasquezNo ratings yet
- Voluntary Eye Movements PDFDocument14 pagesVoluntary Eye Movements PDFsmdiviNo ratings yet
- Mashour2018-Correlates of ConsciousnessDocument3 pagesMashour2018-Correlates of ConsciousnessGiulia AndreeaNo ratings yet
- Internally Generated Reactivation of Single Neurons in Human Hippocampus During Free RecallDocument9 pagesInternally Generated Reactivation of Single Neurons in Human Hippocampus During Free Recalltatiane yoshizawaNo ratings yet
- Study of Neurosecretory Cells in OdonataDocument16 pagesStudy of Neurosecretory Cells in Odonatashubhamatilkar04No ratings yet
- 1 s2.0 S0956566323005808 MainDocument11 pages1 s2.0 S0956566323005808 MainLuís CerdeiraNo ratings yet
- How Does The Brain Organize Information?Document16 pagesHow Does The Brain Organize Information?Fausto Intilla100% (1)
- The Neurobiology of Thought - The Groundbreaking Discoveries of Patricia Goldman Rakic 1937-2003Document13 pagesThe Neurobiology of Thought - The Groundbreaking Discoveries of Patricia Goldman Rakic 1937-2003juan_bacha_1No ratings yet
- Foldable 3D Electrode ArraysDocument14 pagesFoldable 3D Electrode ArraysOrsolya KekesiNo ratings yet
- EEG-Based Perceived Tactile Location Prediction: Deng Wang, Yadong Liu, Dewen Hu, and Gunnar BlohmDocument7 pagesEEG-Based Perceived Tactile Location Prediction: Deng Wang, Yadong Liu, Dewen Hu, and Gunnar BlohmKristianto WibawaNo ratings yet
- Reconstruction of Virtual Neural Circuits in An Insect BrainDocument8 pagesReconstruction of Virtual Neural Circuits in An Insect BrainnithiananthiNo ratings yet
- Dynamic BrainDocument35 pagesDynamic BrainchandusgNo ratings yet
- Expanding The Mirror: Vicarious Activity For Actions, Emotions, and SensationsDocument6 pagesExpanding The Mirror: Vicarious Activity For Actions, Emotions, and SensationsMárcia SoaresNo ratings yet
- Prof. Dr.S.C.Masram Aishwarya D. Atilkar M.SC Zoology (Sem-Iii) Dept. of Zoology RTM University NagpurDocument12 pagesProf. Dr.S.C.Masram Aishwarya D. Atilkar M.SC Zoology (Sem-Iii) Dept. of Zoology RTM University Nagpurshubhamatilkar04No ratings yet
- Pulsed Neural Networks and Their Application: Daniel R. Kunkle Chadd MerriganDocument11 pagesPulsed Neural Networks and Their Application: Daniel R. Kunkle Chadd MerriganinnocentbystanderNo ratings yet
- Total Recall - Using Light To Create and Erase Memories - Science in The NewsDocument7 pagesTotal Recall - Using Light To Create and Erase Memories - Science in The NewsRhod WulfNo ratings yet
- Reconsideración BrocaDocument10 pagesReconsideración BrocaJuan LoperaNo ratings yet
- Explorers of the Black Box: The Search for the Cellular Basis of MemoryFrom EverandExplorers of the Black Box: The Search for the Cellular Basis of MemoryRating: 5 out of 5 stars5/5 (1)
- Chapter 8 Practice TestDocument6 pagesChapter 8 Practice Testcrqin qinNo ratings yet
- Kamus AnatomiDocument42 pagesKamus AnatomiAbdul Al-hadiNo ratings yet
- Biology HSSC 1 Paper IDocument12 pagesBiology HSSC 1 Paper Ikirshnadembra44No ratings yet
- ATAR Human Biology 12 Units 3Document13 pagesATAR Human Biology 12 Units 3FireAwayNo ratings yet
- Urinary SystemDocument5 pagesUrinary SystemJushelle Anne Tigoy Pilare100% (1)
- 3.1 Answers: RememberingDocument13 pages3.1 Answers: RememberingDanny TNo ratings yet
- Pediatric Traumatic Brain Injury: Characteristic Features, Diagnosis, and ManagementDocument12 pagesPediatric Traumatic Brain Injury: Characteristic Features, Diagnosis, and ManagementsylvetriNo ratings yet
- Tropical Fish MedicineDocument5 pagesTropical Fish MedicineCamila Carvalho De SouzaNo ratings yet
- Biochemical Energy Production: Type I Multiple-Choice QuestionsDocument12 pagesBiochemical Energy Production: Type I Multiple-Choice QuestionsKevin Dacatimbang100% (1)
- WIDGB2 Utest Skills 6BDocument2 pagesWIDGB2 Utest Skills 6Brosana guoNo ratings yet
- The Digestive System and Body Metabolism The Digestive System FunctionsDocument23 pagesThe Digestive System and Body Metabolism The Digestive System Functionsvaljulene monteNo ratings yet
- Functional and Molecular Outcomes of The Human Masticatory MusclesDocument14 pagesFunctional and Molecular Outcomes of The Human Masticatory MusclesJyaNo ratings yet
- Electrical Injuries in AnimalsDocument12 pagesElectrical Injuries in AnimalsAndressa PachecoNo ratings yet
- Question PaperDocument19 pagesQuestion PaperSaad NadafNo ratings yet
- Amyloidosis and Gaucher's DiseaseDocument2 pagesAmyloidosis and Gaucher's DiseaseA CNo ratings yet
- Materi Kuliah FK ULM 2022 - Lower Extremity I - Hip & KneeDocument61 pagesMateri Kuliah FK ULM 2022 - Lower Extremity I - Hip & Kneezaki ridhoniNo ratings yet
- VGO 421 ObstetricsDocument159 pagesVGO 421 ObstetricsNaveen BasudeNo ratings yet
- Pain Its Diagnosis and Management in The Rehabilitation of Horses - 2016 - Veterinary Clinics of North America Equine PracticeDocument17 pagesPain Its Diagnosis and Management in The Rehabilitation of Horses - 2016 - Veterinary Clinics of North America Equine PracticeAcupuntura de EquinosNo ratings yet
- Appendix I - Plant TissuesDocument24 pagesAppendix I - Plant TissuesAmeera ChaitramNo ratings yet
- Alleviation of Fertility Disorders Based On Probiotics Induced Immune System Improvement: A ReviewDocument10 pagesAlleviation of Fertility Disorders Based On Probiotics Induced Immune System Improvement: A ReviewDeltaphierhoNo ratings yet
- Amino Sugars by Fazal 1Document16 pagesAmino Sugars by Fazal 1Fazal rahimNo ratings yet
- Dwnload Full Holes Human Anatomy and Physiology 15th Edition Shier Solutions Manual PDFDocument35 pagesDwnload Full Holes Human Anatomy and Physiology 15th Edition Shier Solutions Manual PDFbrecciamoodgflo100% (13)
- (29final) 1125angl FinalDocument133 pages(29final) 1125angl FinalAlok KumarNo ratings yet
- A - Glucagon: Abdulmajeed Rabe3Document56 pagesA - Glucagon: Abdulmajeed Rabe3MONFOLANo ratings yet
- #PRAKTIKUM - TT - 1 - KLAS A - TT - OcDocument4 pages#PRAKTIKUM - TT - 1 - KLAS A - TT - OcDewi MNo ratings yet
- FS 5 Activity Multiple ChoiceDocument5 pagesFS 5 Activity Multiple ChoiceFritzy Gwen BabaNo ratings yet
- Unit:-II Chapter-6. Animal Tissue: Important PointsDocument11 pagesUnit:-II Chapter-6. Animal Tissue: Important PointsChandan KumarNo ratings yet
- Link Download Ebook Anatomy and PhysiologyDocument87 pagesLink Download Ebook Anatomy and Physiologyarif100% (1)
- Combined Science Notes-1Document258 pagesCombined Science Notes-1Premier Car Sales67% (3)
- Experiments On Geotropism An Experiment To Demonstrate That The Root Is Positively Geotropic (Or The Root Shows Positive Geotropism) Apparatus or MaterialsDocument4 pagesExperiments On Geotropism An Experiment To Demonstrate That The Root Is Positively Geotropic (Or The Root Shows Positive Geotropism) Apparatus or MaterialsMutegeki AdrianNo ratings yet
































































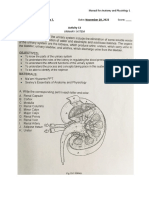

























Download as pdf or txt
You might also like
- Gao Et Al. - 2020 - Neuronal Timescales Are Functionally Dynamic and SDocument26 pagesGao Et Al. - 2020 - Neuronal Timescales Are Functionally Dynamic and Srodrigo carlos carrilloNo ratings yet
- Fernando - Eichenbaum2017Document12 pagesFernando - Eichenbaum2017fernandogfcnsNo ratings yet
- "Inner GPS", A Far-Reaching in Uence in Brain Research-For The Nobel Prize in Physiology or Medicine 2014Document3 pages"Inner GPS", A Far-Reaching in Uence in Brain Research-For The Nobel Prize in Physiology or Medicine 2014mihaela irofteNo ratings yet
- Complexity of Cortical Wave Patterns of The Wake Mouse CortexDocument13 pagesComplexity of Cortical Wave Patterns of The Wake Mouse CortexgrviteNo ratings yet
- Lusk, 2016. Cerebellar Hippocampal and Striatal Time CellsDocument7 pagesLusk, 2016. Cerebellar Hippocampal and Striatal Time CellsAnaMaríaMalagónLNo ratings yet
- A Multi-Modal Parcellation of Human Cerebral Cortex: Europe PMC Funders GroupDocument25 pagesA Multi-Modal Parcellation of Human Cerebral Cortex: Europe PMC Funders GroupYu ZhaoNo ratings yet
- Buzsaki 2004 Nat NeurosciDocument7 pagesBuzsaki 2004 Nat Neuroscihussin elrashidyNo ratings yet
- Tes 111Document2 pagesTes 111Gramy LatumetenNo ratings yet
- Large-Scale Neural Recordings Call For New Insights To Link Brain and BehaviorDocument9 pagesLarge-Scale Neural Recordings Call For New Insights To Link Brain and BehaviorMoshe AlonsoNo ratings yet
- On Time and Space in The Brain - A Relativistic Pseudo-Diffusion FrameworkDocument14 pagesOn Time and Space in The Brain - A Relativistic Pseudo-Diffusion FrameworkGeorge LazarNo ratings yet
- CorrelacionDocument5 pagesCorrelacionlinne25No ratings yet
- Human Premotor Areas Parse Sequences Into Their Spatial and Temporal FeaturesDocument23 pagesHuman Premotor Areas Parse Sequences Into Their Spatial and Temporal FeaturesmoiNo ratings yet
- Neuronal Representation of Saccadic Error in Macaque Posterior Parietal Cortex (PPC)Document17 pagesNeuronal Representation of Saccadic Error in Macaque Posterior Parietal Cortex (PPC)Zeus HeraklesNo ratings yet
- 2015 - Nielson Et Al. - Human Hippocampus Represents Space and Time During Retrieval of Real-World MemoriesDocument6 pages2015 - Nielson Et Al. - Human Hippocampus Represents Space and Time During Retrieval of Real-World MemoriesNityananda PortelladaNo ratings yet
- TMP 4 FB0Document11 pagesTMP 4 FB0FrontiersNo ratings yet
- The Elusive Concept of Brain Connectivity: Barry HorwitzDocument5 pagesThe Elusive Concept of Brain Connectivity: Barry HorwitzParisa SattarNo ratings yet
- Ciliberti Et Al., 2018Document14 pagesCiliberti Et Al., 2018carlos rosalesNo ratings yet
- György Buzsáki - The Brain From Inside Out-Oxford University Press, USA (2019)Document20 pagesGyörgy Buzsáki - The Brain From Inside Out-Oxford University Press, USA (2019)juannnnNo ratings yet
- Paz Gelbard-Sagiv Fried2010-Hippocampal Substrate For Linking Successive EventsDocument6 pagesPaz Gelbard-Sagiv Fried2010-Hippocampal Substrate For Linking Successive Eventsiulia andreeaNo ratings yet
- Theta-Phase Dependent Neuronal Coding During Sequence Learning in Human Single NeuronsDocument12 pagesTheta-Phase Dependent Neuronal Coding During Sequence Learning in Human Single NeuronsWalter HuNo ratings yet
- Journal Pcbi 1011801Document30 pagesJournal Pcbi 1011801shivang patelNo ratings yet
- An Evolving Perspective On The Dynamic Brain - Notes From The Brain Conference On Dynamics of The Brain - Temporal Aspects of ComputationDocument22 pagesAn Evolving Perspective On The Dynamic Brain - Notes From The Brain Conference On Dynamics of The Brain - Temporal Aspects of ComputationRajsuryan SinghNo ratings yet
- Communication From The Cerebellum To The Neocortex During Sleep SpindlesDocument1 pageCommunication From The Cerebellum To The Neocortex During Sleep SpindlesDavid NazarovNo ratings yet
- Temporal EstimationDocument25 pagesTemporal EstimationangieNo ratings yet
- Sereno Et Al., 2020Document6 pagesSereno Et Al., 2020elinNo ratings yet
- Large and Fast Human Pyramidal Neurons Associate With IntelligenceDocument21 pagesLarge and Fast Human Pyramidal Neurons Associate With Intelligenceauf ayeNo ratings yet
- A Cortical Neural Prosthesis For Restoring and Enhancing Memory - Theodore W Berger - Dong Song - Vasilis Z Marmarelis - Et Al - JNeuralEng - 2011 - 1741-2552 - 8!4!046017Document12 pagesA Cortical Neural Prosthesis For Restoring and Enhancing Memory - Theodore W Berger - Dong Song - Vasilis Z Marmarelis - Et Al - JNeuralEng - 2011 - 1741-2552 - 8!4!046017Satyam NegiNo ratings yet
- 174 The Brain From Inside Out 135: Chapter 5. Internalization of ExperienceDocument20 pages174 The Brain From Inside Out 135: Chapter 5. Internalization of ExperiencejuannnnNo ratings yet
- Integrated Neurophotonics Toward Dense Volumetric Interrogation of Brain Circuit Activity-At Depth and in Real Time - 1-s2.0-S0896627320307674-MainDocument2 pagesIntegrated Neurophotonics Toward Dense Volumetric Interrogation of Brain Circuit Activity-At Depth and in Real Time - 1-s2.0-S0896627320307674-MainGeneration GenerationNo ratings yet
- Cognitive Map NobelDocument2 pagesCognitive Map NobelSakditad SaowapaNo ratings yet
- Conectoma Mosca Prepublicación PDFDocument76 pagesConectoma Mosca Prepublicación PDFSofia TombaNo ratings yet
- Using Goal-Driven Deep Learning Models To Understand The Sensory CortexDocument10 pagesUsing Goal-Driven Deep Learning Models To Understand The Sensory CortexRajsuryan SinghNo ratings yet
- Tarantino Et Al 2010Document10 pagesTarantino Et Al 2010jsaccuzzoNo ratings yet
- Review: Conscious Processing and The Global Neuronal Workspace HypothesisDocument23 pagesReview: Conscious Processing and The Global Neuronal Workspace HypothesisSebastian Gonzalez MazoNo ratings yet
- Conscious Visual PerceptionDocument46 pagesConscious Visual PerceptionDa CuNo ratings yet
- Large-Scale Recording of Neuronal Ensembles: PerspectiveDocument6 pagesLarge-Scale Recording of Neuronal Ensembles: PerspectiveMatheus PortelaNo ratings yet
- Sperry Roger Ecs 2003Document7 pagesSperry Roger Ecs 2003Johnmark ObligarNo ratings yet
- Millisecond by Millisecond, Year by Year: Normative EEG Microstates and Developmental StagesDocument5 pagesMillisecond by Millisecond, Year by Year: Normative EEG Microstates and Developmental Stagesscribd.com@jperla.comNo ratings yet
- 1 s2.0 S0092867420310771 MainDocument38 pages1 s2.0 S0092867420310771 MainmyczhNo ratings yet
- Silences Spikes and Bursts Three-Part Knot of TheDocument15 pagesSilences Spikes and Bursts Three-Part Knot of TheUlises ChialvaNo ratings yet
- Mapping Neuronal DiversityDocument2 pagesMapping Neuronal DiversityJohn MistryNo ratings yet
- Simplicial and Topological Descriptions of Human Brain DynamicsDocument20 pagesSimplicial and Topological Descriptions of Human Brain DynamicsJosue VasquezNo ratings yet
- Voluntary Eye Movements PDFDocument14 pagesVoluntary Eye Movements PDFsmdiviNo ratings yet
- Mashour2018-Correlates of ConsciousnessDocument3 pagesMashour2018-Correlates of ConsciousnessGiulia AndreeaNo ratings yet
- Internally Generated Reactivation of Single Neurons in Human Hippocampus During Free RecallDocument9 pagesInternally Generated Reactivation of Single Neurons in Human Hippocampus During Free Recalltatiane yoshizawaNo ratings yet
- Study of Neurosecretory Cells in OdonataDocument16 pagesStudy of Neurosecretory Cells in Odonatashubhamatilkar04No ratings yet
- 1 s2.0 S0956566323005808 MainDocument11 pages1 s2.0 S0956566323005808 MainLuís CerdeiraNo ratings yet
- How Does The Brain Organize Information?Document16 pagesHow Does The Brain Organize Information?Fausto Intilla100% (1)
- The Neurobiology of Thought - The Groundbreaking Discoveries of Patricia Goldman Rakic 1937-2003Document13 pagesThe Neurobiology of Thought - The Groundbreaking Discoveries of Patricia Goldman Rakic 1937-2003juan_bacha_1No ratings yet
- Foldable 3D Electrode ArraysDocument14 pagesFoldable 3D Electrode ArraysOrsolya KekesiNo ratings yet
- EEG-Based Perceived Tactile Location Prediction: Deng Wang, Yadong Liu, Dewen Hu, and Gunnar BlohmDocument7 pagesEEG-Based Perceived Tactile Location Prediction: Deng Wang, Yadong Liu, Dewen Hu, and Gunnar BlohmKristianto WibawaNo ratings yet
- Reconstruction of Virtual Neural Circuits in An Insect BrainDocument8 pagesReconstruction of Virtual Neural Circuits in An Insect BrainnithiananthiNo ratings yet
- Dynamic BrainDocument35 pagesDynamic BrainchandusgNo ratings yet
- Expanding The Mirror: Vicarious Activity For Actions, Emotions, and SensationsDocument6 pagesExpanding The Mirror: Vicarious Activity For Actions, Emotions, and SensationsMárcia SoaresNo ratings yet
- Prof. Dr.S.C.Masram Aishwarya D. Atilkar M.SC Zoology (Sem-Iii) Dept. of Zoology RTM University NagpurDocument12 pagesProf. Dr.S.C.Masram Aishwarya D. Atilkar M.SC Zoology (Sem-Iii) Dept. of Zoology RTM University Nagpurshubhamatilkar04No ratings yet
- Pulsed Neural Networks and Their Application: Daniel R. Kunkle Chadd MerriganDocument11 pagesPulsed Neural Networks and Their Application: Daniel R. Kunkle Chadd MerriganinnocentbystanderNo ratings yet
- Total Recall - Using Light To Create and Erase Memories - Science in The NewsDocument7 pagesTotal Recall - Using Light To Create and Erase Memories - Science in The NewsRhod WulfNo ratings yet
- Reconsideración BrocaDocument10 pagesReconsideración BrocaJuan LoperaNo ratings yet
- Explorers of the Black Box: The Search for the Cellular Basis of MemoryFrom EverandExplorers of the Black Box: The Search for the Cellular Basis of MemoryRating: 5 out of 5 stars5/5 (1)
- Chapter 8 Practice TestDocument6 pagesChapter 8 Practice Testcrqin qinNo ratings yet
- Kamus AnatomiDocument42 pagesKamus AnatomiAbdul Al-hadiNo ratings yet
- Biology HSSC 1 Paper IDocument12 pagesBiology HSSC 1 Paper Ikirshnadembra44No ratings yet
- ATAR Human Biology 12 Units 3Document13 pagesATAR Human Biology 12 Units 3FireAwayNo ratings yet
- Urinary SystemDocument5 pagesUrinary SystemJushelle Anne Tigoy Pilare100% (1)
- 3.1 Answers: RememberingDocument13 pages3.1 Answers: RememberingDanny TNo ratings yet
- Pediatric Traumatic Brain Injury: Characteristic Features, Diagnosis, and ManagementDocument12 pagesPediatric Traumatic Brain Injury: Characteristic Features, Diagnosis, and ManagementsylvetriNo ratings yet
- Tropical Fish MedicineDocument5 pagesTropical Fish MedicineCamila Carvalho De SouzaNo ratings yet
- Biochemical Energy Production: Type I Multiple-Choice QuestionsDocument12 pagesBiochemical Energy Production: Type I Multiple-Choice QuestionsKevin Dacatimbang100% (1)
- WIDGB2 Utest Skills 6BDocument2 pagesWIDGB2 Utest Skills 6Brosana guoNo ratings yet
- The Digestive System and Body Metabolism The Digestive System FunctionsDocument23 pagesThe Digestive System and Body Metabolism The Digestive System Functionsvaljulene monteNo ratings yet
- Functional and Molecular Outcomes of The Human Masticatory MusclesDocument14 pagesFunctional and Molecular Outcomes of The Human Masticatory MusclesJyaNo ratings yet
- Electrical Injuries in AnimalsDocument12 pagesElectrical Injuries in AnimalsAndressa PachecoNo ratings yet
- Question PaperDocument19 pagesQuestion PaperSaad NadafNo ratings yet
- Amyloidosis and Gaucher's DiseaseDocument2 pagesAmyloidosis and Gaucher's DiseaseA CNo ratings yet
- Materi Kuliah FK ULM 2022 - Lower Extremity I - Hip & KneeDocument61 pagesMateri Kuliah FK ULM 2022 - Lower Extremity I - Hip & Kneezaki ridhoniNo ratings yet
- VGO 421 ObstetricsDocument159 pagesVGO 421 ObstetricsNaveen BasudeNo ratings yet
- Pain Its Diagnosis and Management in The Rehabilitation of Horses - 2016 - Veterinary Clinics of North America Equine PracticeDocument17 pagesPain Its Diagnosis and Management in The Rehabilitation of Horses - 2016 - Veterinary Clinics of North America Equine PracticeAcupuntura de EquinosNo ratings yet
- Appendix I - Plant TissuesDocument24 pagesAppendix I - Plant TissuesAmeera ChaitramNo ratings yet
- Alleviation of Fertility Disorders Based On Probiotics Induced Immune System Improvement: A ReviewDocument10 pagesAlleviation of Fertility Disorders Based On Probiotics Induced Immune System Improvement: A ReviewDeltaphierhoNo ratings yet
- Amino Sugars by Fazal 1Document16 pagesAmino Sugars by Fazal 1Fazal rahimNo ratings yet
- Dwnload Full Holes Human Anatomy and Physiology 15th Edition Shier Solutions Manual PDFDocument35 pagesDwnload Full Holes Human Anatomy and Physiology 15th Edition Shier Solutions Manual PDFbrecciamoodgflo100% (13)
- (29final) 1125angl FinalDocument133 pages(29final) 1125angl FinalAlok KumarNo ratings yet
- A - Glucagon: Abdulmajeed Rabe3Document56 pagesA - Glucagon: Abdulmajeed Rabe3MONFOLANo ratings yet
- #PRAKTIKUM - TT - 1 - KLAS A - TT - OcDocument4 pages#PRAKTIKUM - TT - 1 - KLAS A - TT - OcDewi MNo ratings yet
- FS 5 Activity Multiple ChoiceDocument5 pagesFS 5 Activity Multiple ChoiceFritzy Gwen BabaNo ratings yet
- Unit:-II Chapter-6. Animal Tissue: Important PointsDocument11 pagesUnit:-II Chapter-6. Animal Tissue: Important PointsChandan KumarNo ratings yet
- Link Download Ebook Anatomy and PhysiologyDocument87 pagesLink Download Ebook Anatomy and Physiologyarif100% (1)
- Combined Science Notes-1Document258 pagesCombined Science Notes-1Premier Car Sales67% (3)
- Experiments On Geotropism An Experiment To Demonstrate That The Root Is Positively Geotropic (Or The Root Shows Positive Geotropism) Apparatus or MaterialsDocument4 pagesExperiments On Geotropism An Experiment To Demonstrate That The Root Is Positively Geotropic (Or The Root Shows Positive Geotropism) Apparatus or MaterialsMutegeki AdrianNo ratings yet