
Acetylcholine Receptors Key Players in
Acetylcholine Receptors Key Players in
You might also like
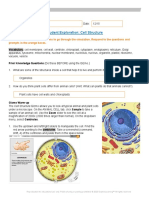
- Cell Structure GizmoDocument4 pagesCell Structure GizmoDavid Yee57% (7)
- Name: Date: Class: Instruction: For Each of The Questions 1 To 50, Choose The Correct Answer For Each Question From The Four Options A, B, C and DDocument7 pagesName: Date: Class: Instruction: For Each of The Questions 1 To 50, Choose The Correct Answer For Each Question From The Four Options A, B, C and Dke2100% (2)
- IAS - Bio - SB1 - Mark Schemes and Assessments PDFDocument74 pagesIAS - Bio - SB1 - Mark Schemes and Assessments PDFisraa jaaboNo ratings yet
- CONCEPT MAP (Central Dogma of Molecular Biology)Document1 pageCONCEPT MAP (Central Dogma of Molecular Biology)ShekinahNo ratings yet
- Regulation of Gastric Electrical and Mechanical Activity by Cholinesterases in MiceDocument17 pagesRegulation of Gastric Electrical and Mechanical Activity by Cholinesterases in MiceRaluca MosoiaNo ratings yet
- AChE y Canceres HumanosDocument66 pagesAChE y Canceres HumanosLuis Antonio FloresNo ratings yet
- AcetylcholineDocument31 pagesAcetylcholineSara KhanNo ratings yet
- PIWI-interacting RNA (piRNA) Signatures in Human Cardiac ProgenitorDocument11 pagesPIWI-interacting RNA (piRNA) Signatures in Human Cardiac Progenitortabrizi.m.fNo ratings yet
- Yoshitoshi-Uebayashi, E. Y. - Modelling Urea-Cycle Disorder Citrullinemia Type 1 With Disease-Specific iPSCs (2017)Document7 pagesYoshitoshi-Uebayashi, E. Y. - Modelling Urea-Cycle Disorder Citrullinemia Type 1 With Disease-Specific iPSCs (2017)FranciscaMesquitaNo ratings yet
- CN 16 338Document12 pagesCN 16 338I Komang Ana MahardikaNo ratings yet
- Acta Histochem 2021 123 5 151717Document7 pagesActa Histochem 2021 123 5 151717Fernando SousaNo ratings yet
- Revi SysteDocument20 pagesRevi Systedr.israquiNo ratings yet
- Ni Hms 479647Document17 pagesNi Hms 479647sirfengchenNo ratings yet
- Alpha7 Nicotinic Acetylcholine Receptor Is A Target in Pharmacology and ToxicologyDocument20 pagesAlpha7 Nicotinic Acetylcholine Receptor Is A Target in Pharmacology and ToxicologyJean Pierre Chastre LuzaNo ratings yet
- Neurochemistry International: Oliver Maier, Julia Böhm, Michael Dahm, Stefan Brück, Cordian Beyer, Sonja JohannDocument10 pagesNeurochemistry International: Oliver Maier, Julia Böhm, Michael Dahm, Stefan Brück, Cordian Beyer, Sonja JohannPili CárdenasNo ratings yet
- Perseverative Behaviour: Crym-Positive Striatal Astrocytes GateDocument42 pagesPerseverative Behaviour: Crym-Positive Striatal Astrocytes Gatejpaulom.No ratings yet
- Journal CancerDocument17 pagesJournal CancerbeklogNo ratings yet
- Extended Atrial Conduction System Characterised by The Expression of The HCN4 Channel and Connexin45Document11 pagesExtended Atrial Conduction System Characterised by The Expression of The HCN4 Channel and Connexin45CARLOS FRANCISCO MANTILLA MONTALVONo ratings yet
- INTRO Rev Neuroseñalización en Tumor de PróstataDocument12 pagesINTRO Rev Neuroseñalización en Tumor de Próstatasarahi venturaNo ratings yet
- Hepatology - 2017 - Han - Circular RNA circMTO1 Acts As The Sponge of microRNA 9 To Suppress Hepatocellular CarcinomaDocument14 pagesHepatology - 2017 - Han - Circular RNA circMTO1 Acts As The Sponge of microRNA 9 To Suppress Hepatocellular CarcinomaSujal PhandNo ratings yet
- Multicentric Cutaneous Neuroendocrine (Merkel Cell) Carcinoma in A DogDocument5 pagesMulticentric Cutaneous Neuroendocrine (Merkel Cell) Carcinoma in A DogPéter VajdovichNo ratings yet
- Ref 6Document12 pagesRef 6CARMEN EMILIA RODRIGUEZ SEMINARIONo ratings yet
- Acetylcholine: Marinar - Picciotto Meenakshialreja Andj - DavidjentschDocument12 pagesAcetylcholine: Marinar - Picciotto Meenakshialreja Andj - DavidjentschAaja SaaudNo ratings yet
- Pulmon Role of Synaptophysin ChromograninDocument9 pagesPulmon Role of Synaptophysin ChromograninJuan Martin HidalgoNo ratings yet
- Nanobiotecnologia: - Nanomedicina - Biomoléculas - BionanoestruturasDocument18 pagesNanobiotecnologia: - Nanomedicina - Biomoléculas - BionanoestruturasAna Margarida MagalhãesNo ratings yet
- 10.1038@s41388 019 1006 0Document13 pages10.1038@s41388 019 1006 0yalocim666No ratings yet
- Tenascin-C Promotes Tumor Cell Migration and Metastasis Through Integrin A9b1-Mediated YAP InhibitionDocument12 pagesTenascin-C Promotes Tumor Cell Migration and Metastasis Through Integrin A9b1-Mediated YAP InhibitionSayda DhaouadiNo ratings yet
- Mast Cells Regulate CD4 T Cell Differentiation 2018 Journal of Allergy andDocument22 pagesMast Cells Regulate CD4 T Cell Differentiation 2018 Journal of Allergy andLuisa FernandaNo ratings yet
- J Otc 2015 11 001Document38 pagesJ Otc 2015 11 001shinaNo ratings yet
- Ajpcell 00166 2015Document22 pagesAjpcell 00166 2015JasonNo ratings yet
- Neuropeptides: Zhi Zhang, Xiang Cao, Xinyu Bao, Yan Zhang, Yun Xu, Dujuan ShaDocument9 pagesNeuropeptides: Zhi Zhang, Xiang Cao, Xinyu Bao, Yan Zhang, Yun Xu, Dujuan ShaIsaac Saucedo FloresNo ratings yet
- 1 s2.0 S1556086416305172 Main PDFDocument10 pages1 s2.0 S1556086416305172 Main PDFlisarahmaasrilNo ratings yet
- Anexina 1Document4 pagesAnexina 1Diana LaviniaNo ratings yet
- MiR 424 Coordinates Multilayered Regulation of Cell Cycle Progres 2018 EBioMDocument12 pagesMiR 424 Coordinates Multilayered Regulation of Cell Cycle Progres 2018 EBioMLuisa FernandaNo ratings yet
- Cells 11 02406Document23 pagesCells 11 02406idboggNo ratings yet
- NIH Public AccessDocument18 pagesNIH Public AccessSofia BottiNo ratings yet
- Acupuncture As Anticancer TreatmentDocument5 pagesAcupuncture As Anticancer TreatmentAlessandro LimaNo ratings yet
- HHS Public Access: Cannabis and Cannabinoid Biology in Stroke: Controversies, Risks, and PromisesDocument13 pagesHHS Public Access: Cannabis and Cannabinoid Biology in Stroke: Controversies, Risks, and PromisesGustavo EspíndolaNo ratings yet
- Atypical Histiocytoid Cells in Metastatic Papillary Thyroid Carcinoma An Underrecognized Cytologic PatternDocument6 pagesAtypical Histiocytoid Cells in Metastatic Papillary Thyroid Carcinoma An Underrecognized Cytologic PatternAnanda FebriantyNo ratings yet
- PC3 Is A Cell Line Characteristic of Prostatic Small Cell CarcinomaDocument12 pagesPC3 Is A Cell Line Characteristic of Prostatic Small Cell CarcinomaANGEL SAUL SERRANO ANGULONo ratings yet
- Medicinal Chemistry Unit III Cholenergic Anti ChokenergicDocument32 pagesMedicinal Chemistry Unit III Cholenergic Anti ChokenergicjalilaNo ratings yet
- EXOSC8Document6 pagesEXOSC8vuhaipham1407No ratings yet
- Case Report: Grace Y. Kim, MD Helen Anaedo, MD Sahar Nozad, MD Tipu Nazeer, MD Hassan Shawa, MDDocument6 pagesCase Report: Grace Y. Kim, MD Helen Anaedo, MD Sahar Nozad, MD Tipu Nazeer, MD Hassan Shawa, MDPedro Gómez RNo ratings yet
- Aptamer and Its Applications in Neurodegenerative Diseases: Cellular and Molecular Life SciencesDocument13 pagesAptamer and Its Applications in Neurodegenerative Diseases: Cellular and Molecular Life SciencesLakshmipriya GopinathNo ratings yet
- 10 1126@scitranslmed Aah6510Document14 pages10 1126@scitranslmed Aah6510翁子貽No ratings yet
- Avances en Neurobiologia de La Union Neuromusuclar Anestesiology 2002Document30 pagesAvances en Neurobiologia de La Union Neuromusuclar Anestesiology 2002Irene ChimalNo ratings yet
- Integrative CUT&Tag-RNA-Seq Analysis of Histone Variant macroH2A1-dependent Orchestration of Human Induced Pluripotent Stem Cell ReprogrammingDocument16 pagesIntegrative CUT&Tag-RNA-Seq Analysis of Histone Variant macroH2A1-dependent Orchestration of Human Induced Pluripotent Stem Cell Reprogrammingniccolo.liorni1No ratings yet
- Lech in New Article 121208Document10 pagesLech in New Article 121208Saidah SaidNo ratings yet
- The Coexistence of Cushing Syndrome and Gynecomast - 2018 - The Kaohsiung JournaDocument2 pagesThe Coexistence of Cushing Syndrome and Gynecomast - 2018 - The Kaohsiung JournaChander YadavNo ratings yet
- Nicotina y ApoptosisDocument17 pagesNicotina y ApoptosisOperacion ConocimientoNo ratings yet
- Expression of A Breast-Cancer-Associated Protein (Ps2) in Human Neuro-Endocrine TumoursDocument5 pagesExpression of A Breast-Cancer-Associated Protein (Ps2) in Human Neuro-Endocrine TumoursSarly FebrianaNo ratings yet
- Dimensions of Neutrophil Life and Fate 2020Document7 pagesDimensions of Neutrophil Life and Fate 2020brunareis1999No ratings yet
- Evaluation of The Susceptibility of Neurons Anticancer DrugsDocument6 pagesEvaluation of The Susceptibility of Neurons Anticancer DrugsLisbeth AlmNo ratings yet
- Astrocytes in Endocannabinoid Signalling: ReviewDocument7 pagesAstrocytes in Endocannabinoid Signalling: ReviewHelington CostaNo ratings yet
- The Place of The Department of Pathological Anatomy of The CHU Ibn Rochd Casablanca in The Diagnosis and Classification of Bronchopulmonary CancerDocument6 pagesThe Place of The Department of Pathological Anatomy of The CHU Ibn Rochd Casablanca in The Diagnosis and Classification of Bronchopulmonary CancerInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- Sinonasal Non-Small Cell Neuroendocrine Carcinoma The Validity of Histological Grading Case Report and A Review of The LiteratureDocument16 pagesSinonasal Non-Small Cell Neuroendocrine Carcinoma The Validity of Histological Grading Case Report and A Review of The Literaturenurul hidayahNo ratings yet
- Jurnal 1 KalsitoninDocument9 pagesJurnal 1 KalsitoninPrawido AndriadiNo ratings yet
- 3 - Mesenchymal Stromal Cells As A Potential Therapeutic For Neurological DisordersDocument6 pages3 - Mesenchymal Stromal Cells As A Potential Therapeutic For Neurological DisordersÊndel AlvesNo ratings yet
- 8-rEPROGRAMMINGDocument16 pages8-rEPROGRAMMINGpaulo saNo ratings yet
- Hu Et Al-2017-Journal of Cellular PhysiologyDocument37 pagesHu Et Al-2017-Journal of Cellular PhysiologySorop AndreiNo ratings yet
- Upregulated Acetylcholine Synthesis During Early Differentiation in The Embryonic Stem Cell Line Cgr8Document5 pagesUpregulated Acetylcholine Synthesis During Early Differentiation in The Embryonic Stem Cell Line Cgr8JasonNo ratings yet
- Stroke and Translational Research - Review of Experimental Models WithDocument17 pagesStroke and Translational Research - Review of Experimental Models WithNST MtzNo ratings yet
- PTX 3Document9 pagesPTX 3M Ali AdrianNo ratings yet
- Clinical Handbook of Cardiac ElectrophysiologyFrom EverandClinical Handbook of Cardiac ElectrophysiologyBenedict M. GloverNo ratings yet
- Protien-Synthesis Lesson PlanDocument8 pagesProtien-Synthesis Lesson PlanHAIDEE VASQUEZNo ratings yet
- Soni Clinic & Pathology Center Chanda: Address:-Front of TVS AgencyDocument1 pageSoni Clinic & Pathology Center Chanda: Address:-Front of TVS AgencyVishalNo ratings yet
- Photosynthesis (Light and Dark Reaction) :: Concept Notes With Formative ActivitiesDocument12 pagesPhotosynthesis (Light and Dark Reaction) :: Concept Notes With Formative ActivitiesMonica SolomonNo ratings yet
- Advantages and Disadvantages of Fermentation and Aerobic RespirationDocument10 pagesAdvantages and Disadvantages of Fermentation and Aerobic RespirationIan Gabriel MayoNo ratings yet
- Cell Type Cell Shape Cell Parts Visible FunctionDocument5 pagesCell Type Cell Shape Cell Parts Visible FunctionAgnesRicardoNo ratings yet
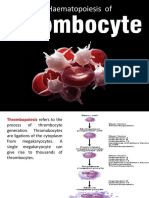
- Trombopoiesis (3) - 1Document10 pagesTrombopoiesis (3) - 1Vannisa Dwi NovianaNo ratings yet
- 12th Zoology Important Creative 235 Mark Questions English MediumDocument63 pages12th Zoology Important Creative 235 Mark Questions English MediumVathi N. Khalleelulah100% (1)
- Gen Bio 1 Love of Lab Group Performance Task 1Document2 pagesGen Bio 1 Love of Lab Group Performance Task 1Ryan GañaNo ratings yet
- Biosensors: Microfluidic-Based Droplets For Advanced Regenerative Medicine: Current Challenges and Future TrendsDocument29 pagesBiosensors: Microfluidic-Based Droplets For Advanced Regenerative Medicine: Current Challenges and Future Trendsmadhu kammarNo ratings yet
- Chapter 3 Micropara (Outline)Document5 pagesChapter 3 Micropara (Outline)Jezrylle BalaongNo ratings yet
- 9 Essential Amino AcidsDocument4 pages9 Essential Amino AcidsimeneNo ratings yet
- Mitosis and MeiosisDocument4 pagesMitosis and MeiosisSanah KhanNo ratings yet
- Unit 1Document10 pagesUnit 1evanjalinNo ratings yet
- Protein Synthesis Essay QuestionsDocument5 pagesProtein Synthesis Essay QuestionsNat Rice100% (1)
- Hematology 1 Mtap1Document70 pagesHematology 1 Mtap1Cyril Llego ManuelNo ratings yet
- Cell Organelles QuizDocument16 pagesCell Organelles Quizعبدالله عفيف الشخصNo ratings yet
- Chapter 10 BloodDocument6 pagesChapter 10 BloodSymonette OcturaNo ratings yet
- Combined HemaDocument28 pagesCombined HemaViena Mae MaglupayNo ratings yet
- Phagocytic Immuno DefficienciesDocument38 pagesPhagocytic Immuno Defficienciesصدام حسینNo ratings yet
- BoichemistryDocument20 pagesBoichemistrynishwa RajpootNo ratings yet
- 2021-2022 S3 AE AnswersDocument12 pages2021-2022 S3 AE AnswersEndi WongNo ratings yet
- Pathology of Skeletal SystemDocument65 pagesPathology of Skeletal SystemFirda S SalsabilaNo ratings yet
- Bio 140 Exam 4 Fall 2015Document7 pagesBio 140 Exam 4 Fall 2015lp_blackoutNo ratings yet
- Plant Tissue Culture of OnionDocument6 pagesPlant Tissue Culture of OnionRegina ChrisantNo ratings yet
- Efficient Mineralization and Osteogenic Gene Overexpression of Mesenchymal Stem Cells On Decellularized Spinach Leaf ScaffoldDocument9 pagesEfficient Mineralization and Osteogenic Gene Overexpression of Mesenchymal Stem Cells On Decellularized Spinach Leaf ScaffoldMartin ContardeNo ratings yet























































































Download as pdf or txt
You might also like
- Cell Structure GizmoDocument4 pagesCell Structure GizmoDavid Yee57% (7)
- Name: Date: Class: Instruction: For Each of The Questions 1 To 50, Choose The Correct Answer For Each Question From The Four Options A, B, C and DDocument7 pagesName: Date: Class: Instruction: For Each of The Questions 1 To 50, Choose The Correct Answer For Each Question From The Four Options A, B, C and Dke2100% (2)
- IAS - Bio - SB1 - Mark Schemes and Assessments PDFDocument74 pagesIAS - Bio - SB1 - Mark Schemes and Assessments PDFisraa jaaboNo ratings yet
- CONCEPT MAP (Central Dogma of Molecular Biology)Document1 pageCONCEPT MAP (Central Dogma of Molecular Biology)ShekinahNo ratings yet
- Regulation of Gastric Electrical and Mechanical Activity by Cholinesterases in MiceDocument17 pagesRegulation of Gastric Electrical and Mechanical Activity by Cholinesterases in MiceRaluca MosoiaNo ratings yet
- AChE y Canceres HumanosDocument66 pagesAChE y Canceres HumanosLuis Antonio FloresNo ratings yet
- AcetylcholineDocument31 pagesAcetylcholineSara KhanNo ratings yet
- PIWI-interacting RNA (piRNA) Signatures in Human Cardiac ProgenitorDocument11 pagesPIWI-interacting RNA (piRNA) Signatures in Human Cardiac Progenitortabrizi.m.fNo ratings yet
- Yoshitoshi-Uebayashi, E. Y. - Modelling Urea-Cycle Disorder Citrullinemia Type 1 With Disease-Specific iPSCs (2017)Document7 pagesYoshitoshi-Uebayashi, E. Y. - Modelling Urea-Cycle Disorder Citrullinemia Type 1 With Disease-Specific iPSCs (2017)FranciscaMesquitaNo ratings yet
- CN 16 338Document12 pagesCN 16 338I Komang Ana MahardikaNo ratings yet
- Acta Histochem 2021 123 5 151717Document7 pagesActa Histochem 2021 123 5 151717Fernando SousaNo ratings yet
- Revi SysteDocument20 pagesRevi Systedr.israquiNo ratings yet
- Ni Hms 479647Document17 pagesNi Hms 479647sirfengchenNo ratings yet
- Alpha7 Nicotinic Acetylcholine Receptor Is A Target in Pharmacology and ToxicologyDocument20 pagesAlpha7 Nicotinic Acetylcholine Receptor Is A Target in Pharmacology and ToxicologyJean Pierre Chastre LuzaNo ratings yet
- Neurochemistry International: Oliver Maier, Julia Böhm, Michael Dahm, Stefan Brück, Cordian Beyer, Sonja JohannDocument10 pagesNeurochemistry International: Oliver Maier, Julia Böhm, Michael Dahm, Stefan Brück, Cordian Beyer, Sonja JohannPili CárdenasNo ratings yet
- Perseverative Behaviour: Crym-Positive Striatal Astrocytes GateDocument42 pagesPerseverative Behaviour: Crym-Positive Striatal Astrocytes Gatejpaulom.No ratings yet
- Journal CancerDocument17 pagesJournal CancerbeklogNo ratings yet
- Extended Atrial Conduction System Characterised by The Expression of The HCN4 Channel and Connexin45Document11 pagesExtended Atrial Conduction System Characterised by The Expression of The HCN4 Channel and Connexin45CARLOS FRANCISCO MANTILLA MONTALVONo ratings yet
- INTRO Rev Neuroseñalización en Tumor de PróstataDocument12 pagesINTRO Rev Neuroseñalización en Tumor de Próstatasarahi venturaNo ratings yet
- Hepatology - 2017 - Han - Circular RNA circMTO1 Acts As The Sponge of microRNA 9 To Suppress Hepatocellular CarcinomaDocument14 pagesHepatology - 2017 - Han - Circular RNA circMTO1 Acts As The Sponge of microRNA 9 To Suppress Hepatocellular CarcinomaSujal PhandNo ratings yet
- Multicentric Cutaneous Neuroendocrine (Merkel Cell) Carcinoma in A DogDocument5 pagesMulticentric Cutaneous Neuroendocrine (Merkel Cell) Carcinoma in A DogPéter VajdovichNo ratings yet
- Ref 6Document12 pagesRef 6CARMEN EMILIA RODRIGUEZ SEMINARIONo ratings yet
- Acetylcholine: Marinar - Picciotto Meenakshialreja Andj - DavidjentschDocument12 pagesAcetylcholine: Marinar - Picciotto Meenakshialreja Andj - DavidjentschAaja SaaudNo ratings yet
- Pulmon Role of Synaptophysin ChromograninDocument9 pagesPulmon Role of Synaptophysin ChromograninJuan Martin HidalgoNo ratings yet
- Nanobiotecnologia: - Nanomedicina - Biomoléculas - BionanoestruturasDocument18 pagesNanobiotecnologia: - Nanomedicina - Biomoléculas - BionanoestruturasAna Margarida MagalhãesNo ratings yet
- 10.1038@s41388 019 1006 0Document13 pages10.1038@s41388 019 1006 0yalocim666No ratings yet
- Tenascin-C Promotes Tumor Cell Migration and Metastasis Through Integrin A9b1-Mediated YAP InhibitionDocument12 pagesTenascin-C Promotes Tumor Cell Migration and Metastasis Through Integrin A9b1-Mediated YAP InhibitionSayda DhaouadiNo ratings yet
- Mast Cells Regulate CD4 T Cell Differentiation 2018 Journal of Allergy andDocument22 pagesMast Cells Regulate CD4 T Cell Differentiation 2018 Journal of Allergy andLuisa FernandaNo ratings yet
- J Otc 2015 11 001Document38 pagesJ Otc 2015 11 001shinaNo ratings yet
- Ajpcell 00166 2015Document22 pagesAjpcell 00166 2015JasonNo ratings yet
- Neuropeptides: Zhi Zhang, Xiang Cao, Xinyu Bao, Yan Zhang, Yun Xu, Dujuan ShaDocument9 pagesNeuropeptides: Zhi Zhang, Xiang Cao, Xinyu Bao, Yan Zhang, Yun Xu, Dujuan ShaIsaac Saucedo FloresNo ratings yet
- 1 s2.0 S1556086416305172 Main PDFDocument10 pages1 s2.0 S1556086416305172 Main PDFlisarahmaasrilNo ratings yet
- Anexina 1Document4 pagesAnexina 1Diana LaviniaNo ratings yet
- MiR 424 Coordinates Multilayered Regulation of Cell Cycle Progres 2018 EBioMDocument12 pagesMiR 424 Coordinates Multilayered Regulation of Cell Cycle Progres 2018 EBioMLuisa FernandaNo ratings yet
- Cells 11 02406Document23 pagesCells 11 02406idboggNo ratings yet
- NIH Public AccessDocument18 pagesNIH Public AccessSofia BottiNo ratings yet
- Acupuncture As Anticancer TreatmentDocument5 pagesAcupuncture As Anticancer TreatmentAlessandro LimaNo ratings yet
- HHS Public Access: Cannabis and Cannabinoid Biology in Stroke: Controversies, Risks, and PromisesDocument13 pagesHHS Public Access: Cannabis and Cannabinoid Biology in Stroke: Controversies, Risks, and PromisesGustavo EspíndolaNo ratings yet
- Atypical Histiocytoid Cells in Metastatic Papillary Thyroid Carcinoma An Underrecognized Cytologic PatternDocument6 pagesAtypical Histiocytoid Cells in Metastatic Papillary Thyroid Carcinoma An Underrecognized Cytologic PatternAnanda FebriantyNo ratings yet
- PC3 Is A Cell Line Characteristic of Prostatic Small Cell CarcinomaDocument12 pagesPC3 Is A Cell Line Characteristic of Prostatic Small Cell CarcinomaANGEL SAUL SERRANO ANGULONo ratings yet
- Medicinal Chemistry Unit III Cholenergic Anti ChokenergicDocument32 pagesMedicinal Chemistry Unit III Cholenergic Anti ChokenergicjalilaNo ratings yet
- EXOSC8Document6 pagesEXOSC8vuhaipham1407No ratings yet
- Case Report: Grace Y. Kim, MD Helen Anaedo, MD Sahar Nozad, MD Tipu Nazeer, MD Hassan Shawa, MDDocument6 pagesCase Report: Grace Y. Kim, MD Helen Anaedo, MD Sahar Nozad, MD Tipu Nazeer, MD Hassan Shawa, MDPedro Gómez RNo ratings yet
- Aptamer and Its Applications in Neurodegenerative Diseases: Cellular and Molecular Life SciencesDocument13 pagesAptamer and Its Applications in Neurodegenerative Diseases: Cellular and Molecular Life SciencesLakshmipriya GopinathNo ratings yet
- 10 1126@scitranslmed Aah6510Document14 pages10 1126@scitranslmed Aah6510翁子貽No ratings yet
- Avances en Neurobiologia de La Union Neuromusuclar Anestesiology 2002Document30 pagesAvances en Neurobiologia de La Union Neuromusuclar Anestesiology 2002Irene ChimalNo ratings yet
- Integrative CUT&Tag-RNA-Seq Analysis of Histone Variant macroH2A1-dependent Orchestration of Human Induced Pluripotent Stem Cell ReprogrammingDocument16 pagesIntegrative CUT&Tag-RNA-Seq Analysis of Histone Variant macroH2A1-dependent Orchestration of Human Induced Pluripotent Stem Cell Reprogrammingniccolo.liorni1No ratings yet
- Lech in New Article 121208Document10 pagesLech in New Article 121208Saidah SaidNo ratings yet
- The Coexistence of Cushing Syndrome and Gynecomast - 2018 - The Kaohsiung JournaDocument2 pagesThe Coexistence of Cushing Syndrome and Gynecomast - 2018 - The Kaohsiung JournaChander YadavNo ratings yet
- Nicotina y ApoptosisDocument17 pagesNicotina y ApoptosisOperacion ConocimientoNo ratings yet
- Expression of A Breast-Cancer-Associated Protein (Ps2) in Human Neuro-Endocrine TumoursDocument5 pagesExpression of A Breast-Cancer-Associated Protein (Ps2) in Human Neuro-Endocrine TumoursSarly FebrianaNo ratings yet
- Dimensions of Neutrophil Life and Fate 2020Document7 pagesDimensions of Neutrophil Life and Fate 2020brunareis1999No ratings yet
- Evaluation of The Susceptibility of Neurons Anticancer DrugsDocument6 pagesEvaluation of The Susceptibility of Neurons Anticancer DrugsLisbeth AlmNo ratings yet
- Astrocytes in Endocannabinoid Signalling: ReviewDocument7 pagesAstrocytes in Endocannabinoid Signalling: ReviewHelington CostaNo ratings yet
- The Place of The Department of Pathological Anatomy of The CHU Ibn Rochd Casablanca in The Diagnosis and Classification of Bronchopulmonary CancerDocument6 pagesThe Place of The Department of Pathological Anatomy of The CHU Ibn Rochd Casablanca in The Diagnosis and Classification of Bronchopulmonary CancerInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- Sinonasal Non-Small Cell Neuroendocrine Carcinoma The Validity of Histological Grading Case Report and A Review of The LiteratureDocument16 pagesSinonasal Non-Small Cell Neuroendocrine Carcinoma The Validity of Histological Grading Case Report and A Review of The Literaturenurul hidayahNo ratings yet
- Jurnal 1 KalsitoninDocument9 pagesJurnal 1 KalsitoninPrawido AndriadiNo ratings yet
- 3 - Mesenchymal Stromal Cells As A Potential Therapeutic For Neurological DisordersDocument6 pages3 - Mesenchymal Stromal Cells As A Potential Therapeutic For Neurological DisordersÊndel AlvesNo ratings yet
- 8-rEPROGRAMMINGDocument16 pages8-rEPROGRAMMINGpaulo saNo ratings yet
- Hu Et Al-2017-Journal of Cellular PhysiologyDocument37 pagesHu Et Al-2017-Journal of Cellular PhysiologySorop AndreiNo ratings yet
- Upregulated Acetylcholine Synthesis During Early Differentiation in The Embryonic Stem Cell Line Cgr8Document5 pagesUpregulated Acetylcholine Synthesis During Early Differentiation in The Embryonic Stem Cell Line Cgr8JasonNo ratings yet
- Stroke and Translational Research - Review of Experimental Models WithDocument17 pagesStroke and Translational Research - Review of Experimental Models WithNST MtzNo ratings yet
- PTX 3Document9 pagesPTX 3M Ali AdrianNo ratings yet
- Clinical Handbook of Cardiac ElectrophysiologyFrom EverandClinical Handbook of Cardiac ElectrophysiologyBenedict M. GloverNo ratings yet
- Protien-Synthesis Lesson PlanDocument8 pagesProtien-Synthesis Lesson PlanHAIDEE VASQUEZNo ratings yet
- Soni Clinic & Pathology Center Chanda: Address:-Front of TVS AgencyDocument1 pageSoni Clinic & Pathology Center Chanda: Address:-Front of TVS AgencyVishalNo ratings yet
- Photosynthesis (Light and Dark Reaction) :: Concept Notes With Formative ActivitiesDocument12 pagesPhotosynthesis (Light and Dark Reaction) :: Concept Notes With Formative ActivitiesMonica SolomonNo ratings yet
- Advantages and Disadvantages of Fermentation and Aerobic RespirationDocument10 pagesAdvantages and Disadvantages of Fermentation and Aerobic RespirationIan Gabriel MayoNo ratings yet
- Cell Type Cell Shape Cell Parts Visible FunctionDocument5 pagesCell Type Cell Shape Cell Parts Visible FunctionAgnesRicardoNo ratings yet
- Trombopoiesis (3) - 1Document10 pagesTrombopoiesis (3) - 1Vannisa Dwi NovianaNo ratings yet
- 12th Zoology Important Creative 235 Mark Questions English MediumDocument63 pages12th Zoology Important Creative 235 Mark Questions English MediumVathi N. Khalleelulah100% (1)
- Gen Bio 1 Love of Lab Group Performance Task 1Document2 pagesGen Bio 1 Love of Lab Group Performance Task 1Ryan GañaNo ratings yet
- Biosensors: Microfluidic-Based Droplets For Advanced Regenerative Medicine: Current Challenges and Future TrendsDocument29 pagesBiosensors: Microfluidic-Based Droplets For Advanced Regenerative Medicine: Current Challenges and Future Trendsmadhu kammarNo ratings yet
- Chapter 3 Micropara (Outline)Document5 pagesChapter 3 Micropara (Outline)Jezrylle BalaongNo ratings yet
- 9 Essential Amino AcidsDocument4 pages9 Essential Amino AcidsimeneNo ratings yet
- Mitosis and MeiosisDocument4 pagesMitosis and MeiosisSanah KhanNo ratings yet
- Unit 1Document10 pagesUnit 1evanjalinNo ratings yet
- Protein Synthesis Essay QuestionsDocument5 pagesProtein Synthesis Essay QuestionsNat Rice100% (1)
- Hematology 1 Mtap1Document70 pagesHematology 1 Mtap1Cyril Llego ManuelNo ratings yet
- Cell Organelles QuizDocument16 pagesCell Organelles Quizعبدالله عفيف الشخصNo ratings yet
- Chapter 10 BloodDocument6 pagesChapter 10 BloodSymonette OcturaNo ratings yet
- Combined HemaDocument28 pagesCombined HemaViena Mae MaglupayNo ratings yet
- Phagocytic Immuno DefficienciesDocument38 pagesPhagocytic Immuno Defficienciesصدام حسینNo ratings yet
- BoichemistryDocument20 pagesBoichemistrynishwa RajpootNo ratings yet
- 2021-2022 S3 AE AnswersDocument12 pages2021-2022 S3 AE AnswersEndi WongNo ratings yet
- Pathology of Skeletal SystemDocument65 pagesPathology of Skeletal SystemFirda S SalsabilaNo ratings yet
- Bio 140 Exam 4 Fall 2015Document7 pagesBio 140 Exam 4 Fall 2015lp_blackoutNo ratings yet
- Plant Tissue Culture of OnionDocument6 pagesPlant Tissue Culture of OnionRegina ChrisantNo ratings yet
- Efficient Mineralization and Osteogenic Gene Overexpression of Mesenchymal Stem Cells On Decellularized Spinach Leaf ScaffoldDocument9 pagesEfficient Mineralization and Osteogenic Gene Overexpression of Mesenchymal Stem Cells On Decellularized Spinach Leaf ScaffoldMartin ContardeNo ratings yet