Download as pdf or txt
You might also like
- Arvophillia Case StudyDocument5 pagesArvophillia Case StudyMickelson Joseph Vil0% (1)
- Bob Beck Protocol - Alternative Cancer TreatmentsDocument15 pagesBob Beck Protocol - Alternative Cancer TreatmentsChristopher Land100% (4)
- The Map Is Still Not The Territory Reflections On Sexual HealthDocument7 pagesThe Map Is Still Not The Territory Reflections On Sexual HealthValerie Walker LagosNo ratings yet
- Review Article Sars Cov-2 and Other Reverse Zoonoses: Understanding The Human Animal Interface For Devising Control StrategiesDocument10 pagesReview Article Sars Cov-2 and Other Reverse Zoonoses: Understanding The Human Animal Interface For Devising Control Strategiescgoswami700No ratings yet
- Elife 66550 v2Document33 pagesElife 66550 v2Anna McMeekinsNo ratings yet
- Clase 1. Evolución de PatogenosDocument30 pagesClase 1. Evolución de PatogenosClaudia Alicia Herrera OlivasNo ratings yet
- Wang 2021Document13 pagesWang 2021Júlia PaulaNo ratings yet
- Impact Defense - Michigan7 2014 GHJPPDocument198 pagesImpact Defense - Michigan7 2014 GHJPPlewa109No ratings yet
- Rylevprevalence and Distribution Pathogen WorldwideDocument16 pagesRylevprevalence and Distribution Pathogen WorldwideazyytieNo ratings yet
- Accepted Manuscript: Acta TropicaDocument49 pagesAccepted Manuscript: Acta TropicaIvonne CabreraNo ratings yet
- Leal Et Al 2000Document8 pagesLeal Et Al 2000Luciana SaundersNo ratings yet
- Biodiversity Inhibits Parasites: Broad Evidence For The Dilution EffectDocument6 pagesBiodiversity Inhibits Parasites: Broad Evidence For The Dilution EffectMuziNo ratings yet
- Role of Vertically Transmitted Viral and Bacterial EndosymbiontsDocument15 pagesRole of Vertically Transmitted Viral and Bacterial EndosymbiontsAfraNo ratings yet
- International Journal For Parasitology: A B B C C D D e F G H I B J KDocument10 pagesInternational Journal For Parasitology: A B B C C D D e F G H I B J KViviana GeraldinNo ratings yet
- Research Paper On ZoonosesDocument8 pagesResearch Paper On Zoonosesaflbtvmfp100% (1)
- Dónde Se Presentará La Nueva PandemiaDocument8 pagesDónde Se Presentará La Nueva PandemiajuanvillicanaNo ratings yet
- First Epidemiological Survey On The Prevalence and Subtypes Distribution of The Enteric Parasite Blastocystis Sp. in VietnamDocument11 pagesFirst Epidemiological Survey On The Prevalence and Subtypes Distribution of The Enteric Parasite Blastocystis Sp. in VietnamDavid A. Osorio G.No ratings yet
- Peerj 12272Document25 pagesPeerj 12272Camila Mercedes Trujillo FigueroaNo ratings yet
- Evolução Da Resistência em Mosquitos (2018)Document14 pagesEvolução Da Resistência em Mosquitos (2018)Gilberto Muniz JúniorNo ratings yet
- Fmicb 11 631736Document16 pagesFmicb 11 631736Herick Lyncon Antunes RodriguesNo ratings yet
- Kerteszia Cruzii and Extra-Amazonian Malaria in Brazil Challenges Due To Climate Change in The Atlantic ForestDocument10 pagesKerteszia Cruzii and Extra-Amazonian Malaria in Brazil Challenges Due To Climate Change in The Atlantic ForestThiago AzevedoNo ratings yet
- Journal of Animal Ecology - 2016 - Clark - Co Infections and Environmental Conditions Drive The Distributions of BloodDocument10 pagesJournal of Animal Ecology - 2016 - Clark - Co Infections and Environmental Conditions Drive The Distributions of BloodGustavo AndresNo ratings yet
- Delator Re 2021Document14 pagesDelator Re 2021Gustavo AndresNo ratings yet
- Arbovirus-Mosquito Vector-Host Interactions and The Impact On Transmission and Disease Pathogenesis of ArbovirusesDocument14 pagesArbovirus-Mosquito Vector-Host Interactions and The Impact On Transmission and Disease Pathogenesis of ArbovirusesLúcia ReisNo ratings yet
- New - Epidemiological Infectious DiseasesDocument25 pagesNew - Epidemiological Infectious DiseasesSharmaine MararacNo ratings yet
- BEERNTSEN Et Al 2000Document23 pagesBEERNTSEN Et Al 2000Ana Beatriz Vasquez RodriguezNo ratings yet
- Mordecai - 2013 - Consequences of Pathogen Spillover For Cheatgrass-Invaded Grasslands Coexistence, Competitive Exclusion, or Priority E-AnnotatedDocument11 pagesMordecai - 2013 - Consequences of Pathogen Spillover For Cheatgrass-Invaded Grasslands Coexistence, Competitive Exclusion, or Priority E-AnnotatedrikriknlNo ratings yet
- Huang Et Al 2021 Infectious Disease Macroecology Parasite Diversity and Dynamics Across The GlobeDocument8 pagesHuang Et Al 2021 Infectious Disease Macroecology Parasite Diversity and Dynamics Across The GlobeanurofaunaNo ratings yet
- Lung Flukes Eco-FitDocument9 pagesLung Flukes Eco-FitSondang FarwatyNo ratings yet
- Design and Validation of A Novel Multi-Epitopes Vaccine Against HantavirusDocument12 pagesDesign and Validation of A Novel Multi-Epitopes Vaccine Against HantavirusOgunleke AdebolaNo ratings yet
- Giardia Duodenalis in Patients With Diarrhea and Various Animals in Northeastern ChinaDocument11 pagesGiardia Duodenalis in Patients With Diarrhea and Various Animals in Northeastern ChinaFlávia UchôaNo ratings yet
- The Quest For A True One Health Perspective of BrucellosisDocument18 pagesThe Quest For A True One Health Perspective of Brucellosisyogui1965No ratings yet
- Fgene 13 918227Document12 pagesFgene 13 918227BarbaraNo ratings yet
- Global Rise in Human Infectious DiseaseDocument6 pagesGlobal Rise in Human Infectious DiseaseViona GustianaNo ratings yet
- Zoonotic Tuberculosis: A Review of Epidemiology, Clinical Presentation, Prevention and ControlDocument7 pagesZoonotic Tuberculosis: A Review of Epidemiology, Clinical Presentation, Prevention and ControlGigih Rahmadani MarisNo ratings yet
- ArtigosDocument7 pagesArtigosLeoberto TorresNo ratings yet
- Demographic and Genetic FactorsgenderDocument8 pagesDemographic and Genetic FactorsgenderIsabella CarbonariNo ratings yet
- Host Specificity, Pathogen Exposure, and Superinfections Impact TheDocument14 pagesHost Specificity, Pathogen Exposure, and Superinfections Impact TheSarai MontielNo ratings yet
- Epi1 IntroDocument16 pagesEpi1 IntroKAUSHIK GUPTANo ratings yet
- Influenza VirusDocument2 pagesInfluenza VirusSeele TlhagaNo ratings yet
- Foodborne Parasitic Diseases in The Neotropics A ReviewDocument15 pagesFoodborne Parasitic Diseases in The Neotropics A Reviewestefasalazar2419No ratings yet
- Jvi 01778-22Document14 pagesJvi 01778-22Paulo BorattoNo ratings yet
- AscarisDocument26 pagesAscarisLaboratorio EmergenciaNo ratings yet
- Climate Change and Vector-Borne Diseases of Public Health SignificanceDocument8 pagesClimate Change and Vector-Borne Diseases of Public Health SignificanceAriadne Cristina De Antonio100% (1)
- Pandemics and Global WarningDocument3 pagesPandemics and Global Warningalessandro.crivellaroNo ratings yet
- 573-Article Text-3611-1-10-20220117Document10 pages573-Article Text-3611-1-10-20220117vivilanciaNo ratings yet
- vms3 434Document9 pagesvms3 434ali sherNo ratings yet
- Ni Hms 811004Document20 pagesNi Hms 811004cristina_gomhNo ratings yet
- Biosafety and Health: SciencedirectDocument7 pagesBiosafety and Health: SciencedirectIslam Ausraf RajibNo ratings yet
- Vega Heredia2011Document11 pagesVega Heredia2011andiniNo ratings yet
- Altizer Et Al. - 2011 - Animal Migration and Infectious Disease RiskDocument7 pagesAltizer Et Al. - 2011 - Animal Migration and Infectious Disease RiskbahanbugNo ratings yet
- Infcetious Disease Ecology of Wild BirdsDocument278 pagesInfcetious Disease Ecology of Wild BirdsVarunNo ratings yet
- Human Fascioliasis Infection Sources Their Diversity Incidence Factors Analytical Methods and Prevention Measures PDFDocument35 pagesHuman Fascioliasis Infection Sources Their Diversity Incidence Factors Analytical Methods and Prevention Measures PDFmuchlis syahnuddinNo ratings yet
- 1 s2.0 S0048969720361040 MainDocument8 pages1 s2.0 S0048969720361040 MainWemerson DanielNo ratings yet
- 01 AEXTJ 389 23 RevisedDocument6 pages01 AEXTJ 389 23 RevisedBRNSS Publication Hub InfoNo ratings yet
- Epidemiology of Vector-Borne DiseasesDocument17 pagesEpidemiology of Vector-Borne Diseaseselizabeth cortesNo ratings yet
- Co-Circulation of in Uenza A Virus Strains and Emergence of Pandemic Via Reassortment: The Role of Cross-ImmunityDocument14 pagesCo-Circulation of in Uenza A Virus Strains and Emergence of Pandemic Via Reassortment: The Role of Cross-ImmunityAn TaraNo ratings yet
- Leprosy in FLORIDA, USA - 231215 - 023236Document3 pagesLeprosy in FLORIDA, USA - 231215 - 023236dr samyNo ratings yet
- Evolución Viral en La Emergencia de ZoonosisDocument23 pagesEvolución Viral en La Emergencia de ZoonosisCarlos MendozaNo ratings yet
- Annan 2023Document8 pagesAnnan 2023Ana Beatriz Vasquez RodriguezNo ratings yet
- Zoonoticdis 03 00014Document3 pagesZoonoticdis 03 00014Jelena KilićNo ratings yet
- Orthohantavirus Pathogenesis and Cell TropismDocument14 pagesOrthohantavirus Pathogenesis and Cell TropismJesus Barraza SánchezNo ratings yet
- Apocalypse Now: Global Collapse and the Virus Threat: Global Collapse, #4From EverandApocalypse Now: Global Collapse and the Virus Threat: Global Collapse, #4No ratings yet
- Mental Health Wellness Trainer in Albuquerque NM Resume Carol BruscaDocument3 pagesMental Health Wellness Trainer in Albuquerque NM Resume Carol BruscaCarolBruscaNo ratings yet
- Cutaneous T.BDocument60 pagesCutaneous T.BSajin AlexanderNo ratings yet
- Diagnostic Protocol ModificatDocument83 pagesDiagnostic Protocol Modificatuda20130% (1)
- Intrapartum Care For Healthy Women and Babies PDF 35109866447557Document96 pagesIntrapartum Care For Healthy Women and Babies PDF 35109866447557Lakshmi DheviNo ratings yet
- Requests For MED3 Fit Note': Covid-19 Guidance For PracticesDocument2 pagesRequests For MED3 Fit Note': Covid-19 Guidance For Practicesplayjake18No ratings yet
- Acute Sinusitis: Practice EssentialsDocument22 pagesAcute Sinusitis: Practice EssentialsAna-Maria DuMiNo ratings yet
- Disease and Astrology PDFDocument9 pagesDisease and Astrology PDFAbhishek P BenjaminNo ratings yet
- Qualities of An Officiating OfficialsDocument3 pagesQualities of An Officiating OfficialsMark Anthony Estoque Dusal80% (5)
- Antibiotic Induced Black Hairy TongueDocument11 pagesAntibiotic Induced Black Hairy TongueYuganya SriNo ratings yet
- All in One Homeopathic Materia Medica Niranjan Mohanty.01209 2Document6 pagesAll in One Homeopathic Materia Medica Niranjan Mohanty.01209 2ballaramesh1968No ratings yet
- Ca4657en PDFDocument1 pageCa4657en PDFAndres ValdezNo ratings yet
- Testicular BiopsyDocument110 pagesTesticular Biopsyabhinav_choudhary03No ratings yet
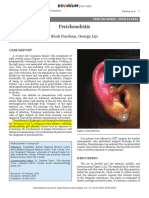
- Perichondritis: Shah Darshan, George LijiDocument4 pagesPerichondritis: Shah Darshan, George LijiThasia IsabelitaNo ratings yet
- Properties of Skeletal MuscleDocument72 pagesProperties of Skeletal MuscleAakash GohilNo ratings yet
- Mapeh 10 Quarter 4 NotesDocument5 pagesMapeh 10 Quarter 4 NotesJohrence ReyesNo ratings yet
- Grape Seed & Skin Extract As Potential PrebioticDocument14 pagesGrape Seed & Skin Extract As Potential PrebioticSébastien Le QuéréNo ratings yet
- Mydev Module 5 Answer SheetDocument4 pagesMydev Module 5 Answer SheetAtlas ParejaNo ratings yet
- Promoting Care For Child Development in Community Health ServicesDocument34 pagesPromoting Care For Child Development in Community Health ServicesMaria Kurchatova Lopes PereiraNo ratings yet
- ML Minor MayDocument5 pagesML Minor Maygovind kumarNo ratings yet
- UltraMedicRider Takaful ENG PDFDocument2 pagesUltraMedicRider Takaful ENG PDFDhyra Osem ScienceNo ratings yet
- Chapter 3 3 - Nutritional Assessment Methods Biochemical Clinical - PDFDocument36 pagesChapter 3 3 - Nutritional Assessment Methods Biochemical Clinical - PDFDegefu TadesseNo ratings yet
- Ferrous Sulfate: o o o o o o oDocument5 pagesFerrous Sulfate: o o o o o o oLelanie Japitana100% (1)
- Fucidin H Cream: Fusidic Acid and Hydrocortisone AcetateDocument2 pagesFucidin H Cream: Fusidic Acid and Hydrocortisone Acetateسید طاہر شیرازیNo ratings yet
- Malaria: DR Mohammedyassin RediDocument63 pagesMalaria: DR Mohammedyassin Redimiki amareNo ratings yet
- 24 May 22 GNLMDocument15 pages24 May 22 GNLMlaiNo ratings yet
- Patniyot Irene Headache in AdolescentsDocument16 pagesPatniyot Irene Headache in AdolescentsAnonymous tG35SYROzENo ratings yet
- Vision & Mission: Healthc Are Solution GroupDocument16 pagesVision & Mission: Healthc Are Solution GroupFajar Pandu WijayaNo ratings yet