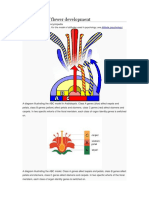
Download as pdf or txt
You might also like
- Development: Flowering TransitionDocument3 pagesDevelopment: Flowering TransitionNovember RolfNo ratings yet
- Development: InflorescenceDocument3 pagesDevelopment: InflorescenceArjel JamiasNo ratings yet
- tmp684 TMPDocument6 pagestmp684 TMPFrontiersNo ratings yet
- Flower DevelopmentDocument17 pagesFlower DevelopmentNurlinaNo ratings yet
- Flower DevelopmentDocument12 pagesFlower DevelopmentthenderingNo ratings yet
- Plcell v1 1 37Document16 pagesPlcell v1 1 37HaienasrNo ratings yet
- CH 24 PDFDocument32 pagesCH 24 PDFANGIE DANIELA PRADA PLATANo ratings yet
- MBU Genetic Control of FlowersDocument4 pagesMBU Genetic Control of FlowersketakeeNo ratings yet
- ABC Model of Flower DevelopmentDocument11 pagesABC Model of Flower DevelopmentAlexandra Maria NeaguNo ratings yet
- Bunga 4Document2 pagesBunga 4yosephineikaaNo ratings yet
- BIO109P Module 2. Differentiation & MorphogenesisDocument28 pagesBIO109P Module 2. Differentiation & MorphogenesisJet DelimaNo ratings yet
- Unit 9Document38 pagesUnit 9Sejal PrasadNo ratings yet
- The Roles of Plant Hormones and Their Interactions With Regulatory Genes in Determining Meristem ActivityDocument19 pagesThe Roles of Plant Hormones and Their Interactions With Regulatory Genes in Determining Meristem Activitymohamed rouaniNo ratings yet
- ABC Model of Flower Development - WikipediaDocument9 pagesABC Model of Flower Development - WikipediaCarina JLNo ratings yet
- SAM Charles 2003Document8 pagesSAM Charles 2003AdriandiNo ratings yet
- Molecular Mechanisms of Flower Development: An Armchair GuideDocument11 pagesMolecular Mechanisms of Flower Development: An Armchair GuideCarina JLNo ratings yet
- Molecular Genetic Approaches To Plant Development ReviewDocument16 pagesMolecular Genetic Approaches To Plant Development ReviewFirman M. NurNo ratings yet
- Flower Induction and Flower Bud Development in Apple and SweetDocument11 pagesFlower Induction and Flower Bud Development in Apple and Sweetssllppl48No ratings yet
- Inflorescence CyperaceaeInflorDocument21 pagesInflorescence CyperaceaeInflorGhulamJelaniNo ratings yet
- Genetics of Development in Plants - NOTESDocument40 pagesGenetics of Development in Plants - NOTESSanthoshNo ratings yet
- Stamen Structure and Function: R. J. Scott, M. Spielman, and H. G. DickinsonDocument15 pagesStamen Structure and Function: R. J. Scott, M. Spielman, and H. G. DickinsonAJMRNo ratings yet
- Introduction To Plant Science Flower and Inflorescence StructureDocument17 pagesIntroduction To Plant Science Flower and Inflorescence StructureAlan ReidNo ratings yet
- HDocument2 pagesHLevi GraceNo ratings yet
- Flower: BlossomDocument22 pagesFlower: BlossomEmpire kingNo ratings yet
- Lec 1 CRPDocument26 pagesLec 1 CRPavddorajeeraoNo ratings yet
- The Making of The Ower: July 2005Document7 pagesThe Making of The Ower: July 2005BerryNo ratings yet
- Functional Roles of Histone Modification, Chromatin Remodeling and Micrornas in Arabidopsis Flower DevelopmentDocument47 pagesFunctional Roles of Histone Modification, Chromatin Remodeling and Micrornas in Arabidopsis Flower DevelopmentZain BaderNo ratings yet
- Exercise 8 DevbiolabDocument9 pagesExercise 8 DevbiolabPrezyNo ratings yet
- Developmental Plant BiogicalDocument30 pagesDevelopmental Plant BiogicalJuan Manuel Colorado LaraNo ratings yet
- Sugars and Flowering in The Grapevine (Vitis Vinifera L.)Document14 pagesSugars and Flowering in The Grapevine (Vitis Vinifera L.)Deivid Silva de SouzaNo ratings yet
- Artikel BungaDocument2 pagesArtikel BunganenesNo ratings yet
- Biology Organisms and Adaptations Media Update Enhanced Edition 1st Edition Noyd Solutions Manual DownloadDocument22 pagesBiology Organisms and Adaptations Media Update Enhanced Edition 1st Edition Noyd Solutions Manual DownloadKaren Zavala100% (30)
- Exercise 10Document4 pagesExercise 10drsabuasNo ratings yet
- Reproduction in Plants: 8.1 F - A F O ADocument22 pagesReproduction in Plants: 8.1 F - A F O Atechnically whoNo ratings yet
- Flower: Jump To Navigation Jump To Search Flower (Disambiguation) Floral (Disambiguation)Document12 pagesFlower: Jump To Navigation Jump To Search Flower (Disambiguation) Floral (Disambiguation)jolinmartinNo ratings yet
- Structural Organisation in Plants and Animals: Rationalised 2023-24Document16 pagesStructural Organisation in Plants and Animals: Rationalised 2023-24Mohamed Fayaz Sheik DawoodNo ratings yet
- What Makes Adventitious RootsDocument24 pagesWhat Makes Adventitious RootsEdersin TorresNo ratings yet
- Flower: From Wikipedia, The Free EncyclopediaDocument14 pagesFlower: From Wikipedia, The Free EncyclopediaBibithaNo ratings yet
- Floracion VidDocument11 pagesFloracion VidCamiloNo ratings yet
- Leaf Development in DicotsDocument6 pagesLeaf Development in DicotsVish BhakNo ratings yet
- Agr 516 - Laboratory ManualDocument20 pagesAgr 516 - Laboratory ManualAiman WafiyNo ratings yet
- Morphology: Floral PartsDocument8 pagesMorphology: Floral PartsKiana DouglasNo ratings yet
- FlowerDocument22 pagesFlowerGrety IancuNo ratings yet
- Morphology of Plant NCERTDocument21 pagesMorphology of Plant NCERTnishukr5454No ratings yet
- Flower: Navigation Search Flower (Disambiguation) Floral (Disambiguation)Document18 pagesFlower: Navigation Search Flower (Disambiguation) Floral (Disambiguation)devakikris1985No ratings yet
- Inflorescence, Pollination and FertilizationDocument16 pagesInflorescence, Pollination and Fertilizationniswati zahroNo ratings yet
- 12th Class Biology Ncert Chap-2Document23 pages12th Class Biology Ncert Chap-2Prathyusha MannemNo ratings yet
- Taxonomic Evidence: Structural and Biochemical Characters: MorphologyDocument48 pagesTaxonomic Evidence: Structural and Biochemical Characters: MorphologyGhulamJelaniNo ratings yet
- Goethe, Sex, and Flower Genes: Letter To The EditorDocument4 pagesGoethe, Sex, and Flower Genes: Letter To The EditorVish BhakNo ratings yet
- Sex DeterminationDocument11 pagesSex DeterminationJulio SubagioNo ratings yet
- Mechanism of FloweringDocument40 pagesMechanism of FloweringIrdatul WardahNo ratings yet
- Phyllot in GerberaDocument11 pagesPhyllot in GerberaStevo JobosNo ratings yet
- Dendrology 1Document17 pagesDendrology 1Jone BlushNo ratings yet
- Senses and Signals Review Current ScienceDocument10 pagesSenses and Signals Review Current ScienceNiranjana Prasanna VenkateswaranNo ratings yet
- Morphology in Flowering PlantsDocument39 pagesMorphology in Flowering Plantscoolakshat3664No ratings yet
- Haywood S303 25Document24 pagesHaywood S303 25Mart KarmNo ratings yet
- Flower: From Wikipedia, The Free EncyclopediaDocument13 pagesFlower: From Wikipedia, The Free EncyclopediaRakesh KumarNo ratings yet
- Das 2016Document38 pagesDas 2016rismaNo ratings yet
- Selina Solutions For Class 9 Biology Chapter 6 Seeds Structure and GerminationDocument8 pagesSelina Solutions For Class 9 Biology Chapter 6 Seeds Structure and GerminationSagnik MajumderNo ratings yet
- Plants ClassificationDocument11 pagesPlants ClassificationLeslie GandaNo ratings yet
- Grading Experiment BiologyDocument3 pagesGrading Experiment BiologyAnaRosuNo ratings yet
- 1a BIO 002 Lecture, KadiriDocument27 pages1a BIO 002 Lecture, KadiriAikay EmmaNo ratings yet
- Lab Report: Departent of Agriculture and Agribusness Management Vegetable and Flower Seed ProductionDocument8 pagesLab Report: Departent of Agriculture and Agribusness Management Vegetable and Flower Seed ProductionAkif KhanNo ratings yet
- Bio CH6 F5 StudywithadminDocument7 pagesBio CH6 F5 StudywithadminRathi MalarNo ratings yet
- Lesson Plan Morphology of Flowering PlantsDocument12 pagesLesson Plan Morphology of Flowering PlantsSHILPA AGARWAL100% (1)
- Plant Classification DegreeDocument57 pagesPlant Classification Degreejohnsonsamuel090677746No ratings yet
- Sample PDF of Neet Ug Biology Book Vol 1Document36 pagesSample PDF of Neet Ug Biology Book Vol 1Luciefer 0035No ratings yet
- Review NotesDocument4 pagesReview NoteshonnyremotoNo ratings yet
- Un Estudio de TargioniaDocument13 pagesUn Estudio de TargioniaramongonzaNo ratings yet
- P5 Science Learner's BookDocument112 pagesP5 Science Learner's BookIt’s me AudreyNo ratings yet
- Tax Morfo Ast Cardueae Susanna2019Document11 pagesTax Morfo Ast Cardueae Susanna2019J CamiloNo ratings yet
- Morphology of Flowering PlantsDocument13 pagesMorphology of Flowering Plantschiragbhaskar16No ratings yet
- Activity 11 - The Leaf (Instruction Sheet)Document9 pagesActivity 11 - The Leaf (Instruction Sheet)Chris P. BeaconNo ratings yet
- Neotropical PhyllanthusDocument88 pagesNeotropical PhyllanthusRayaneRibeiroNo ratings yet
- Seed Germination Inquiry Pre-Inquiry InvestigationDocument19 pagesSeed Germination Inquiry Pre-Inquiry InvestigationjeetuckNo ratings yet
- Plant PropagationDocument36 pagesPlant PropagationDeepa ChoudharyNo ratings yet
- The SeedDocument4 pagesThe SeedAnonymous fe9UIiNo ratings yet
- 3 0 Plant PhysiologyDocument61 pages3 0 Plant Physiologyapi-309062221No ratings yet
- Double FertilizationDocument4 pagesDouble FertilizationHemanta JenaNo ratings yet
- Parts of A FlowerDocument26 pagesParts of A Flowerapi-243961057No ratings yet
- PlantsDocument5 pagesPlantsShapee ManzanitasNo ratings yet
- THE Cross - Section of A Tree: Prepared By: S. BryanDocument17 pagesTHE Cross - Section of A Tree: Prepared By: S. BryanBereket BlessingNo ratings yet
- II. Seed-GerminationDocument49 pagesII. Seed-GerminationAndrew GatesNo ratings yet
- Morphology PremedDocument50 pagesMorphology PremedAASHISHNo ratings yet
- Plant Morphology - DaunDocument50 pagesPlant Morphology - DaunOlik SholikhahNo ratings yet
- Science - IDocument160 pagesScience - IRanuka Hewa-AmarappuligeNo ratings yet
- Growth Developt Pearl MilletDocument17 pagesGrowth Developt Pearl MilletdarmaNo ratings yet
- AngiospermsDocument11 pagesAngiospermstj_simodlanNo ratings yet