
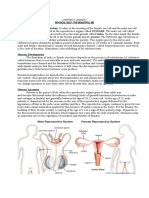
Ashley - REPRODUCTIVE DEVELOPMENT OF THE FEMALE
Ashley - REPRODUCTIVE DEVELOPMENT OF THE FEMALE
You might also like
- The Veil - Cascade PDFDocument232 pagesThe Veil - Cascade PDFSaul100% (4)
- Chapter 19: Audit of Owners' Equity: Review QuestionsDocument18 pagesChapter 19: Audit of Owners' Equity: Review QuestionsReznakNo ratings yet
- Investigatory Project of BiologyDocument18 pagesInvestigatory Project of BiologyRanjana Singh89% (35)
- Social Networking Problems Among Uitm Shah Alam StudentsDocument21 pagesSocial Networking Problems Among Uitm Shah Alam StudentsCik Tiem Ngagiman100% (1)
- Emergency Evacuation Drill ScenarioDocument5 pagesEmergency Evacuation Drill ScenarioiMaaz RameyNo ratings yet
- Clinical Assessment of Child and Adolescent Intelligence - Randy W Kamphaus PDFDocument685 pagesClinical Assessment of Child and Adolescent Intelligence - Randy W Kamphaus PDFCamilo Suárez100% (1)
- Feedback Part of A System (Dylan Wiliam)Document4 pagesFeedback Part of A System (Dylan Wiliam)ajmccarthynzNo ratings yet
- Sarong, MyrelDocument2 pagesSarong, MyrelMyrel Kay SarongNo ratings yet
- Gender and Society - Modules 2 & 3Document17 pagesGender and Society - Modules 2 & 3Lexter JonNo ratings yet
- S10Q3M2 - The Menstrual Cycle PDFDocument46 pagesS10Q3M2 - The Menstrual Cycle PDFLorainne CasnoNo ratings yet
- DuittozDocument21 pagesDuittozJoana FreitasNo ratings yet
- Reproductive SystemDocument11 pagesReproductive SystemnbcNo ratings yet
- Bab IiDocument15 pagesBab IiTinukNo ratings yet
- First Page PDFDocument1 pageFirst Page PDFAndrian SirghiNo ratings yet
- The Normal Menstrual Cycle in WomenDocument4 pagesThe Normal Menstrual Cycle in Womenmartinaloreyn.delrosarioNo ratings yet
- Unitive and Procreative Health Procreation, Creation and EvolutionDocument9 pagesUnitive and Procreative Health Procreation, Creation and EvolutionJustJ ThingsNo ratings yet
- BookDocument33 pagesBookObiogwu MichaelNo ratings yet
- Fertilization Reflection PaperDocument2 pagesFertilization Reflection PaperCrisandro Allen Lazo100% (2)
- REPRODUCTIVEDocument12 pagesREPRODUCTIVEBalolot RalphNo ratings yet
- Fertilitas Dan Infertilitas: BY Tetti Solehati, S.KP., M.KepDocument32 pagesFertilitas Dan Infertilitas: BY Tetti Solehati, S.KP., M.KepWindy Shintia FandinyNo ratings yet
- Fisiologi Reproduksi Kel.6FISIOLOGI REPRODUKSIDocument14 pagesFisiologi Reproduksi Kel.6FISIOLOGI REPRODUKSItiaraarisenda100% (1)
- Expo 2 Science (Physiology)Document9 pagesExpo 2 Science (Physiology)Ale RegaladoNo ratings yet
- INFERTILITYDocument10 pagesINFERTILITYSakshi SangleNo ratings yet
- Unit-2 (13) Lifespan Psychology ModDocument10 pagesUnit-2 (13) Lifespan Psychology Modashish1981No ratings yet
- Stages of Puberty - Sundhed - DKDocument6 pagesStages of Puberty - Sundhed - DKPavel BerlinschiNo ratings yet
- Human Reproductive SystemDocument4 pagesHuman Reproductive SystemJoseph CristianNo ratings yet
- R&S Health Presentation1Document24 pagesR&S Health Presentation1Nidhi KaushikNo ratings yet
- Worksheet Q3 Week 2Document2 pagesWorksheet Q3 Week 2Jaybie TejadaNo ratings yet
- Fertilitas Dan Infertilitas: BY Tettisolehati, S.KP., M.KepDocument32 pagesFertilitas Dan Infertilitas: BY Tettisolehati, S.KP., M.KepAny AndrianiNo ratings yet
- Unit 2 Summary NotesDocument14 pagesUnit 2 Summary NotesQuesyNo ratings yet
- Chapter 7: Sexual Self: - Understanding Basic of Sexual BehaviorDocument12 pagesChapter 7: Sexual Self: - Understanding Basic of Sexual BehaviorJanderick DeveraNo ratings yet
- MODULE 1-Lesson 2-3Document11 pagesMODULE 1-Lesson 2-3JOHN BRAINARD PEJONo ratings yet
- Unit 2 by SuryaDocument29 pagesUnit 2 by SuryayashiranjNo ratings yet
- Gender MODULE 3Document18 pagesGender MODULE 3Raquel Venancio WaklinNo ratings yet
- Chapter 2 Unpacking The Self The Sexual SelfDocument3 pagesChapter 2 Unpacking The Self The Sexual SelfcondrillondenmarjohnNo ratings yet
- Sexual SelfDocument13 pagesSexual SelfswitbbgirlNo ratings yet
- Examining The Number of Births A Woman (Or A Man) Has Produced by A Given Point in Time, Such As The Date of A Census or SurveyDocument17 pagesExamining The Number of Births A Woman (Or A Man) Has Produced by A Given Point in Time, Such As The Date of A Census or SurveyMayjane MicayasNo ratings yet
- ReproductionDocument42 pagesReproductionValentina Bermudez CadavidNo ratings yet
- Pediatric UrologyDocument14 pagesPediatric UrologycibunNo ratings yet
- Module 2 Gender Society 1Document32 pagesModule 2 Gender Society 1John Syvel QuintroNo ratings yet
- Reproduction and Population Control: Module - 3Document26 pagesReproduction and Population Control: Module - 3Jessa Ariño MoralesNo ratings yet
- Meyer Et Al. 2004Document15 pagesMeyer Et Al. 2004Aline Freitas de MeloNo ratings yet
- Physiologic Events of Embryo Implantation and Decidualization in Human and Non-Human PrimatesDocument21 pagesPhysiologic Events of Embryo Implantation and Decidualization in Human and Non-Human PrimatesAnh Vũ Hồ Ngọc100% (1)
- The Nursing Role in Reproductive and Sexual Health: Learning OutcomesDocument11 pagesThe Nursing Role in Reproductive and Sexual Health: Learning OutcomesEric GanzanNo ratings yet
- LESSON 8: Sexual SelfDocument6 pagesLESSON 8: Sexual SelfJeya Plays YTNo ratings yet
- Understanding The Self Chapter 2: Unpacking The Self Miss KZ Ulboc REFERENCE: Cierva, M Chapter 2: Lesson 1 Physical Self: The Beautiful MeDocument5 pagesUnderstanding The Self Chapter 2: Unpacking The Self Miss KZ Ulboc REFERENCE: Cierva, M Chapter 2: Lesson 1 Physical Self: The Beautiful MeKimNo ratings yet
- MODULE 4 - Week 2 - The Child and Adolescent Learners and Learning PrinciplesDocument7 pagesMODULE 4 - Week 2 - The Child and Adolescent Learners and Learning PrinciplesMarsha MGNo ratings yet
- Diagnosis of PregnancyDocument28 pagesDiagnosis of PregnancySuel'n StpdNo ratings yet
- Review Ontology and Endocrinology of The Reproductive System of Bulls From Fetus To MaturityDocument8 pagesReview Ontology and Endocrinology of The Reproductive System of Bulls From Fetus To MaturitySyahzidane YusufNo ratings yet
- Menstrual Cycle: Onset and FrequencyDocument7 pagesMenstrual Cycle: Onset and FrequencyAlloiBialbaNo ratings yet
- Lague - Case Analysis ReportDocument43 pagesLague - Case Analysis Reportinah krizia lagueNo ratings yet
- Ovo LationDocument5 pagesOvo LationAsif Hasan NiloyNo ratings yet
- Goats Reproduction Estrual Cycle Estrous Synchronization Artificial InseminationDocument11 pagesGoats Reproduction Estrual Cycle Estrous Synchronization Artificial InseminationBayu Anke MardianNo ratings yet
- Unit 2 PDFDocument10 pagesUnit 2 PDFRajeshNo ratings yet
- Human Reproductive System - Wikipedia PDFDocument4 pagesHuman Reproductive System - Wikipedia PDFAubrey EuropeNo ratings yet
- The Impact of Age and BMI On Oocyte Maturation and Embryo DevelopmentDocument14 pagesThe Impact of Age and BMI On Oocyte Maturation and Embryo DevelopmentFarah S KadhimNo ratings yet
- Reproductive Physiology and Anatomy of The Sow PDFDocument4 pagesReproductive Physiology and Anatomy of The Sow PDFCarol OrequesNo ratings yet
- Human Life History: Postnatal DevelopmentDocument49 pagesHuman Life History: Postnatal DevelopmentrobiahanitaNo ratings yet
- Ovulation: Eman Star QadrDocument9 pagesOvulation: Eman Star QadrDlzar AbubakrNo ratings yet
- The Process of ReproductionDocument4 pagesThe Process of ReproductionIvan BixenmanNo ratings yet
- Exercise and The Menstrual CycleDocument14 pagesExercise and The Menstrual CycleAndrew HaslettNo ratings yet
- Hubungan Pengetahuan Tentang Menstruasi Dengan Sikap Siswi Kelas Vii Tentang Kebersihan Diri Saat MenstruasiDocument8 pagesHubungan Pengetahuan Tentang Menstruasi Dengan Sikap Siswi Kelas Vii Tentang Kebersihan Diri Saat MenstruasiUlfvaNo ratings yet
- Reading Module I Lesson IIIDocument12 pagesReading Module I Lesson IIIElla Nika FangonNo ratings yet
- Reproductive Cycles in MammalsDocument9 pagesReproductive Cycles in MammalsSeemal shahidNo ratings yet
- Initiation: See AlsoDocument2 pagesInitiation: See AlsoPelícula RoblesNo ratings yet
- Ep English Teachers GuideDocument180 pagesEp English Teachers GuideJessy ChrisNo ratings yet
- 921-Article Text-3249-1-10-20220601Document14 pages921-Article Text-3249-1-10-20220601YuliaNo ratings yet
- TDS 202 Dura ProofDocument2 pagesTDS 202 Dura ProofGhulam WaheedNo ratings yet
- Date Name Age Sex PT Numberward DiagnosisDocument177 pagesDate Name Age Sex PT Numberward Diagnosiskibira peterNo ratings yet
- Brief Hydrogeological Studies of Watershed MR-03 (37) in Context of Groundwater Estimation, Washi, Osmanabad, Maharashtra, IndiaDocument13 pagesBrief Hydrogeological Studies of Watershed MR-03 (37) in Context of Groundwater Estimation, Washi, Osmanabad, Maharashtra, IndiaInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- Math4190 Lecture-04-02 HDocument11 pagesMath4190 Lecture-04-02 Hchandan.thakurNo ratings yet
- Karoora 2012 L-3 (2013)Document9 pagesKaroora 2012 L-3 (2013)FiraolNo ratings yet
- An Overview of Croatian Autochthonous Varieties of Sweet CherryDocument7 pagesAn Overview of Croatian Autochthonous Varieties of Sweet CherryJasna HasanbegovićNo ratings yet
- Development of 22-kV Distribution Systems and Switchgear: Shuichi Kikukawa Kenji Tsuchiya Satoru Kajiwara Akira TakahamaDocument7 pagesDevelopment of 22-kV Distribution Systems and Switchgear: Shuichi Kikukawa Kenji Tsuchiya Satoru Kajiwara Akira TakahamaDave ChaudhuryNo ratings yet
- Iep AidenDocument2 pagesIep Aidenapi-554127143No ratings yet
- NEWS ARTICLE and SHOT LIST-Head of ATMIS Joins Ugandan Nationals in Somalia To Mark 61st Independence DayDocument5 pagesNEWS ARTICLE and SHOT LIST-Head of ATMIS Joins Ugandan Nationals in Somalia To Mark 61st Independence DayAMISOM Public Information ServicesNo ratings yet
- Belt Conveyour Safetyv2Document24 pagesBelt Conveyour Safetyv2Luis MecNo ratings yet
- Ra Sapnotes Res DWLDDocument24 pagesRa Sapnotes Res DWLDramanarekha7206No ratings yet
- Champschicken Com Seocheck 2020 12 17Document17 pagesChampschicken Com Seocheck 2020 12 17Dayanand baraNo ratings yet
- 2020a1t182 Assgn1Document9 pages2020a1t182 Assgn1Aahib NazirNo ratings yet
- Maze ProblemDocument2 pagesMaze ProblemBhuvaneswari RamamurthyNo ratings yet
- The Impact of Food Branding On Children's Eating Behaviour and ObesityDocument8 pagesThe Impact of Food Branding On Children's Eating Behaviour and ObesityAlessandraBattagliaNo ratings yet
- Service ManualDocument283 pagesService ManualcoquerasNo ratings yet
- Présentation XanLite 2020 ENDocument18 pagesPrésentation XanLite 2020 ENJ.DoeNo ratings yet
- Active Directory GP Programming and Registry ReferenceDocument99 pagesActive Directory GP Programming and Registry ReferenceHarmandeep Singh SagguNo ratings yet
- List - Parts of Bahay Na Bato - Filipiniana 101Document7 pagesList - Parts of Bahay Na Bato - Filipiniana 101Eriellynn Liza100% (1)
- Build Underwater RobotDocument3 pagesBuild Underwater RobotSid VjNo ratings yet
- TPM Mean Autonomous MaintenanceDocument27 pagesTPM Mean Autonomous Maintenancesydeng100% (1)























































































Download as docx, pdf, or txt
You might also like
- The Veil - Cascade PDFDocument232 pagesThe Veil - Cascade PDFSaul100% (4)
- Chapter 19: Audit of Owners' Equity: Review QuestionsDocument18 pagesChapter 19: Audit of Owners' Equity: Review QuestionsReznakNo ratings yet
- Investigatory Project of BiologyDocument18 pagesInvestigatory Project of BiologyRanjana Singh89% (35)
- Social Networking Problems Among Uitm Shah Alam StudentsDocument21 pagesSocial Networking Problems Among Uitm Shah Alam StudentsCik Tiem Ngagiman100% (1)
- Emergency Evacuation Drill ScenarioDocument5 pagesEmergency Evacuation Drill ScenarioiMaaz RameyNo ratings yet
- Clinical Assessment of Child and Adolescent Intelligence - Randy W Kamphaus PDFDocument685 pagesClinical Assessment of Child and Adolescent Intelligence - Randy W Kamphaus PDFCamilo Suárez100% (1)
- Feedback Part of A System (Dylan Wiliam)Document4 pagesFeedback Part of A System (Dylan Wiliam)ajmccarthynzNo ratings yet
- Sarong, MyrelDocument2 pagesSarong, MyrelMyrel Kay SarongNo ratings yet
- Gender and Society - Modules 2 & 3Document17 pagesGender and Society - Modules 2 & 3Lexter JonNo ratings yet
- S10Q3M2 - The Menstrual Cycle PDFDocument46 pagesS10Q3M2 - The Menstrual Cycle PDFLorainne CasnoNo ratings yet
- DuittozDocument21 pagesDuittozJoana FreitasNo ratings yet
- Reproductive SystemDocument11 pagesReproductive SystemnbcNo ratings yet
- Bab IiDocument15 pagesBab IiTinukNo ratings yet
- First Page PDFDocument1 pageFirst Page PDFAndrian SirghiNo ratings yet
- The Normal Menstrual Cycle in WomenDocument4 pagesThe Normal Menstrual Cycle in Womenmartinaloreyn.delrosarioNo ratings yet
- Unitive and Procreative Health Procreation, Creation and EvolutionDocument9 pagesUnitive and Procreative Health Procreation, Creation and EvolutionJustJ ThingsNo ratings yet
- BookDocument33 pagesBookObiogwu MichaelNo ratings yet
- Fertilization Reflection PaperDocument2 pagesFertilization Reflection PaperCrisandro Allen Lazo100% (2)
- REPRODUCTIVEDocument12 pagesREPRODUCTIVEBalolot RalphNo ratings yet
- Fertilitas Dan Infertilitas: BY Tetti Solehati, S.KP., M.KepDocument32 pagesFertilitas Dan Infertilitas: BY Tetti Solehati, S.KP., M.KepWindy Shintia FandinyNo ratings yet
- Fisiologi Reproduksi Kel.6FISIOLOGI REPRODUKSIDocument14 pagesFisiologi Reproduksi Kel.6FISIOLOGI REPRODUKSItiaraarisenda100% (1)
- Expo 2 Science (Physiology)Document9 pagesExpo 2 Science (Physiology)Ale RegaladoNo ratings yet
- INFERTILITYDocument10 pagesINFERTILITYSakshi SangleNo ratings yet
- Unit-2 (13) Lifespan Psychology ModDocument10 pagesUnit-2 (13) Lifespan Psychology Modashish1981No ratings yet
- Stages of Puberty - Sundhed - DKDocument6 pagesStages of Puberty - Sundhed - DKPavel BerlinschiNo ratings yet
- Human Reproductive SystemDocument4 pagesHuman Reproductive SystemJoseph CristianNo ratings yet
- R&S Health Presentation1Document24 pagesR&S Health Presentation1Nidhi KaushikNo ratings yet
- Worksheet Q3 Week 2Document2 pagesWorksheet Q3 Week 2Jaybie TejadaNo ratings yet
- Fertilitas Dan Infertilitas: BY Tettisolehati, S.KP., M.KepDocument32 pagesFertilitas Dan Infertilitas: BY Tettisolehati, S.KP., M.KepAny AndrianiNo ratings yet
- Unit 2 Summary NotesDocument14 pagesUnit 2 Summary NotesQuesyNo ratings yet
- Chapter 7: Sexual Self: - Understanding Basic of Sexual BehaviorDocument12 pagesChapter 7: Sexual Self: - Understanding Basic of Sexual BehaviorJanderick DeveraNo ratings yet
- MODULE 1-Lesson 2-3Document11 pagesMODULE 1-Lesson 2-3JOHN BRAINARD PEJONo ratings yet
- Unit 2 by SuryaDocument29 pagesUnit 2 by SuryayashiranjNo ratings yet
- Gender MODULE 3Document18 pagesGender MODULE 3Raquel Venancio WaklinNo ratings yet
- Chapter 2 Unpacking The Self The Sexual SelfDocument3 pagesChapter 2 Unpacking The Self The Sexual SelfcondrillondenmarjohnNo ratings yet
- Sexual SelfDocument13 pagesSexual SelfswitbbgirlNo ratings yet
- Examining The Number of Births A Woman (Or A Man) Has Produced by A Given Point in Time, Such As The Date of A Census or SurveyDocument17 pagesExamining The Number of Births A Woman (Or A Man) Has Produced by A Given Point in Time, Such As The Date of A Census or SurveyMayjane MicayasNo ratings yet
- ReproductionDocument42 pagesReproductionValentina Bermudez CadavidNo ratings yet
- Pediatric UrologyDocument14 pagesPediatric UrologycibunNo ratings yet
- Module 2 Gender Society 1Document32 pagesModule 2 Gender Society 1John Syvel QuintroNo ratings yet
- Reproduction and Population Control: Module - 3Document26 pagesReproduction and Population Control: Module - 3Jessa Ariño MoralesNo ratings yet
- Meyer Et Al. 2004Document15 pagesMeyer Et Al. 2004Aline Freitas de MeloNo ratings yet
- Physiologic Events of Embryo Implantation and Decidualization in Human and Non-Human PrimatesDocument21 pagesPhysiologic Events of Embryo Implantation and Decidualization in Human and Non-Human PrimatesAnh Vũ Hồ Ngọc100% (1)
- The Nursing Role in Reproductive and Sexual Health: Learning OutcomesDocument11 pagesThe Nursing Role in Reproductive and Sexual Health: Learning OutcomesEric GanzanNo ratings yet
- LESSON 8: Sexual SelfDocument6 pagesLESSON 8: Sexual SelfJeya Plays YTNo ratings yet
- Understanding The Self Chapter 2: Unpacking The Self Miss KZ Ulboc REFERENCE: Cierva, M Chapter 2: Lesson 1 Physical Self: The Beautiful MeDocument5 pagesUnderstanding The Self Chapter 2: Unpacking The Self Miss KZ Ulboc REFERENCE: Cierva, M Chapter 2: Lesson 1 Physical Self: The Beautiful MeKimNo ratings yet
- MODULE 4 - Week 2 - The Child and Adolescent Learners and Learning PrinciplesDocument7 pagesMODULE 4 - Week 2 - The Child and Adolescent Learners and Learning PrinciplesMarsha MGNo ratings yet
- Diagnosis of PregnancyDocument28 pagesDiagnosis of PregnancySuel'n StpdNo ratings yet
- Review Ontology and Endocrinology of The Reproductive System of Bulls From Fetus To MaturityDocument8 pagesReview Ontology and Endocrinology of The Reproductive System of Bulls From Fetus To MaturitySyahzidane YusufNo ratings yet
- Menstrual Cycle: Onset and FrequencyDocument7 pagesMenstrual Cycle: Onset and FrequencyAlloiBialbaNo ratings yet
- Lague - Case Analysis ReportDocument43 pagesLague - Case Analysis Reportinah krizia lagueNo ratings yet
- Ovo LationDocument5 pagesOvo LationAsif Hasan NiloyNo ratings yet
- Goats Reproduction Estrual Cycle Estrous Synchronization Artificial InseminationDocument11 pagesGoats Reproduction Estrual Cycle Estrous Synchronization Artificial InseminationBayu Anke MardianNo ratings yet
- Unit 2 PDFDocument10 pagesUnit 2 PDFRajeshNo ratings yet
- Human Reproductive System - Wikipedia PDFDocument4 pagesHuman Reproductive System - Wikipedia PDFAubrey EuropeNo ratings yet
- The Impact of Age and BMI On Oocyte Maturation and Embryo DevelopmentDocument14 pagesThe Impact of Age and BMI On Oocyte Maturation and Embryo DevelopmentFarah S KadhimNo ratings yet
- Reproductive Physiology and Anatomy of The Sow PDFDocument4 pagesReproductive Physiology and Anatomy of The Sow PDFCarol OrequesNo ratings yet
- Human Life History: Postnatal DevelopmentDocument49 pagesHuman Life History: Postnatal DevelopmentrobiahanitaNo ratings yet
- Ovulation: Eman Star QadrDocument9 pagesOvulation: Eman Star QadrDlzar AbubakrNo ratings yet
- The Process of ReproductionDocument4 pagesThe Process of ReproductionIvan BixenmanNo ratings yet
- Exercise and The Menstrual CycleDocument14 pagesExercise and The Menstrual CycleAndrew HaslettNo ratings yet
- Hubungan Pengetahuan Tentang Menstruasi Dengan Sikap Siswi Kelas Vii Tentang Kebersihan Diri Saat MenstruasiDocument8 pagesHubungan Pengetahuan Tentang Menstruasi Dengan Sikap Siswi Kelas Vii Tentang Kebersihan Diri Saat MenstruasiUlfvaNo ratings yet
- Reading Module I Lesson IIIDocument12 pagesReading Module I Lesson IIIElla Nika FangonNo ratings yet
- Reproductive Cycles in MammalsDocument9 pagesReproductive Cycles in MammalsSeemal shahidNo ratings yet
- Initiation: See AlsoDocument2 pagesInitiation: See AlsoPelícula RoblesNo ratings yet
- Ep English Teachers GuideDocument180 pagesEp English Teachers GuideJessy ChrisNo ratings yet
- 921-Article Text-3249-1-10-20220601Document14 pages921-Article Text-3249-1-10-20220601YuliaNo ratings yet
- TDS 202 Dura ProofDocument2 pagesTDS 202 Dura ProofGhulam WaheedNo ratings yet
- Date Name Age Sex PT Numberward DiagnosisDocument177 pagesDate Name Age Sex PT Numberward Diagnosiskibira peterNo ratings yet
- Brief Hydrogeological Studies of Watershed MR-03 (37) in Context of Groundwater Estimation, Washi, Osmanabad, Maharashtra, IndiaDocument13 pagesBrief Hydrogeological Studies of Watershed MR-03 (37) in Context of Groundwater Estimation, Washi, Osmanabad, Maharashtra, IndiaInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- Math4190 Lecture-04-02 HDocument11 pagesMath4190 Lecture-04-02 Hchandan.thakurNo ratings yet
- Karoora 2012 L-3 (2013)Document9 pagesKaroora 2012 L-3 (2013)FiraolNo ratings yet
- An Overview of Croatian Autochthonous Varieties of Sweet CherryDocument7 pagesAn Overview of Croatian Autochthonous Varieties of Sweet CherryJasna HasanbegovićNo ratings yet
- Development of 22-kV Distribution Systems and Switchgear: Shuichi Kikukawa Kenji Tsuchiya Satoru Kajiwara Akira TakahamaDocument7 pagesDevelopment of 22-kV Distribution Systems and Switchgear: Shuichi Kikukawa Kenji Tsuchiya Satoru Kajiwara Akira TakahamaDave ChaudhuryNo ratings yet
- Iep AidenDocument2 pagesIep Aidenapi-554127143No ratings yet
- NEWS ARTICLE and SHOT LIST-Head of ATMIS Joins Ugandan Nationals in Somalia To Mark 61st Independence DayDocument5 pagesNEWS ARTICLE and SHOT LIST-Head of ATMIS Joins Ugandan Nationals in Somalia To Mark 61st Independence DayAMISOM Public Information ServicesNo ratings yet
- Belt Conveyour Safetyv2Document24 pagesBelt Conveyour Safetyv2Luis MecNo ratings yet
- Ra Sapnotes Res DWLDDocument24 pagesRa Sapnotes Res DWLDramanarekha7206No ratings yet
- Champschicken Com Seocheck 2020 12 17Document17 pagesChampschicken Com Seocheck 2020 12 17Dayanand baraNo ratings yet
- 2020a1t182 Assgn1Document9 pages2020a1t182 Assgn1Aahib NazirNo ratings yet
- Maze ProblemDocument2 pagesMaze ProblemBhuvaneswari RamamurthyNo ratings yet
- The Impact of Food Branding On Children's Eating Behaviour and ObesityDocument8 pagesThe Impact of Food Branding On Children's Eating Behaviour and ObesityAlessandraBattagliaNo ratings yet
- Service ManualDocument283 pagesService ManualcoquerasNo ratings yet
- Présentation XanLite 2020 ENDocument18 pagesPrésentation XanLite 2020 ENJ.DoeNo ratings yet
- Active Directory GP Programming and Registry ReferenceDocument99 pagesActive Directory GP Programming and Registry ReferenceHarmandeep Singh SagguNo ratings yet
- List - Parts of Bahay Na Bato - Filipiniana 101Document7 pagesList - Parts of Bahay Na Bato - Filipiniana 101Eriellynn Liza100% (1)
- Build Underwater RobotDocument3 pagesBuild Underwater RobotSid VjNo ratings yet
- TPM Mean Autonomous MaintenanceDocument27 pagesTPM Mean Autonomous Maintenancesydeng100% (1)