Download as pdf or txt
You might also like
- Narrative Report On Smea: Division of Cebu Province Rosalino L. Arreglado Elementary SchoolDocument2 pagesNarrative Report On Smea: Division of Cebu Province Rosalino L. Arreglado Elementary SchoolAngel Gelig100% (17)
- Reading: Healthy Lifestyle Role For All Society'Document4 pagesReading: Healthy Lifestyle Role For All Society'Tore CusimanoNo ratings yet
- Suppressive Effect of Normal and Autoimmune: of Hyperbaric Oxygenation MiceDocument6 pagesSuppressive Effect of Normal and Autoimmune: of Hyperbaric Oxygenation MiceDahlia della KurniaNo ratings yet
- TMP A915Document17 pagesTMP A915FrontiersNo ratings yet
- Primary Immune System Responders To Nucleus Pulposus Cells: Evidence For Immune Response in Disc HerniationDocument9 pagesPrimary Immune System Responders To Nucleus Pulposus Cells: Evidence For Immune Response in Disc HerniationSari Yanti MatsyahNo ratings yet
- 1990 - Recombinant Human Interferon-Y - Differences in Glycosylation and Proteolytic Processing Lead To Heterogeneity in Batch CultureDocument5 pages1990 - Recombinant Human Interferon-Y - Differences in Glycosylation and Proteolytic Processing Lead To Heterogeneity in Batch Culture顏世隆No ratings yet
- Immunology00256 0071Document10 pagesImmunology00256 0071carlitoseverectNo ratings yet
- Measurement of Tumor Necrosis Factor Activity Flow CytometryDocument4 pagesMeasurement of Tumor Necrosis Factor Activity Flow CytometryHendroNo ratings yet
- Veterinary Immunology and ImmunopathologyDocument7 pagesVeterinary Immunology and Immunopathologyjunior5787No ratings yet
- Impaired Cutaneous Immune Responses in Thy-1-De Cient Mice: Related ContentDocument8 pagesImpaired Cutaneous Immune Responses in Thy-1-De Cient Mice: Related ContentcarlitoseverectNo ratings yet
- Cytokine Gene Expression in Skin of Susceptible Guinea-Pig Infected WithDocument6 pagesCytokine Gene Expression in Skin of Susceptible Guinea-Pig Infected Withpasid harlisaNo ratings yet
- Protective Roles of Mast Cells Against Enterobacterial Infection Are Mediated by Toll-Like Receptor 4Document7 pagesProtective Roles of Mast Cells Against Enterobacterial Infection Are Mediated by Toll-Like Receptor 4harumiaikoNo ratings yet
- Ac MonoclonalDocument7 pagesAc MonoclonalRobert Larry Cordova DiazNo ratings yet
- Chapman 1950Document5 pagesChapman 1950poojajain86No ratings yet
- TNF Alpha Is A Key Mediator of Gut Inflammation Seen in Amebic Colitis in Human Intestine in The SCID Mouse-Human Intestinal Xenograft ModelDocument8 pagesTNF Alpha Is A Key Mediator of Gut Inflammation Seen in Amebic Colitis in Human Intestine in The SCID Mouse-Human Intestinal Xenograft Modelu77No ratings yet
- 1241 Full PDFDocument13 pages1241 Full PDFMuhammad FaheemNo ratings yet
- mjm4911 1003Document8 pagesmjm4911 1003Aleja CollazosNo ratings yet
- 1 s2.0 S0034528896901137 MainDocument7 pages1 s2.0 S0034528896901137 MainEdu AlmeidaNo ratings yet
- Immunohistochemical Characterization of A Renal Nephroblastoma in A Trp53-Mutant and Prolyl Isomerase 1-Deficient MouseDocument5 pagesImmunohistochemical Characterization of A Renal Nephroblastoma in A Trp53-Mutant and Prolyl Isomerase 1-Deficient Mouselily rogaNo ratings yet
- Autocrine Interleukin-Lp Regulates Both Proliferation and Apoptosis inDocument6 pagesAutocrine Interleukin-Lp Regulates Both Proliferation and Apoptosis inMauricio LinaresNo ratings yet
- tmp7286 TMPDocument15 pagestmp7286 TMPFrontiersNo ratings yet
- tmp337B TMPDocument29 pagestmp337B TMPFrontiersNo ratings yet
- Eosinophils Protect Nematode Parasite Larvae by Regulating Local ImmunityDocument20 pagesEosinophils Protect Nematode Parasite Larvae by Regulating Local ImmunityLeonardo DonatiNo ratings yet
- II 1101006633Document10 pagesII 1101006633Rodrigo AlvesNo ratings yet
- Mitake Mushroom 3Document6 pagesMitake Mushroom 3dayra maría ixa marín salamancaNo ratings yet
- Nothing To DoDocument11 pagesNothing To DoDiana Muela MoraNo ratings yet
- Immunomodulation in The Canine Endometrium by UteropathogenicDocument17 pagesImmunomodulation in The Canine Endometrium by UteropathogenicAndreea Blanco VeraNo ratings yet
- 377Document14 pages377زيكو زيكوNo ratings yet
- Animal Models of Human DiseaseDocument9 pagesAnimal Models of Human DiseaseamamggNo ratings yet
- Melendres Chem-103 Final-Requirements May-2022Document31 pagesMelendres Chem-103 Final-Requirements May-2022Bianca MelendresNo ratings yet
- SESIÃ"N 9 - Ciclo CelularDocument6 pagesSESIÃ"N 9 - Ciclo CelularIsaac LoyolaNo ratings yet
- Effect of Human and Murine Interferon-A On Steroid Production by Rat Ovarian CellsDocument12 pagesEffect of Human and Murine Interferon-A On Steroid Production by Rat Ovarian CellsJosué VelázquezNo ratings yet
- In Vitro Inhibitory Effects of Antioxidants On Cytotoxicity of T-2 ToxinDocument6 pagesIn Vitro Inhibitory Effects of Antioxidants On Cytotoxicity of T-2 ToxinElijah James LegaspiNo ratings yet
- En Haemonchus TextoDocument6 pagesEn Haemonchus TextoJuliana GattiniNo ratings yet
- Metabolic Control of Progenitor Cell Propagation During Drosophila Tracheal RemodelingDocument17 pagesMetabolic Control of Progenitor Cell Propagation During Drosophila Tracheal Remodelingdede chenNo ratings yet
- Cytokine: Haiqi He, Kenneth J. Genovese, Michael H. KogutDocument7 pagesCytokine: Haiqi He, Kenneth J. Genovese, Michael H. KogutChetan Vilas JawaleNo ratings yet
- Lund 2006Document17 pagesLund 2006marialuizacdasilvaNo ratings yet
- Fujioka 1999Document9 pagesFujioka 1999Sharon ChiangNo ratings yet
- TMP 1 B46Document8 pagesTMP 1 B46FrontiersNo ratings yet
- Apoptosis in Natural Rabies Virus Infection in DogsDocument5 pagesApoptosis in Natural Rabies Virus Infection in DogspapiipiiNo ratings yet
- Interleukin-10 Gene Therapy-Mediated Amelioration of Bacterial PneumoniaDocument7 pagesInterleukin-10 Gene Therapy-Mediated Amelioration of Bacterial Pneumoniaaniekfk95No ratings yet
- Laquinimod EAE 1 Vez DíaDocument10 pagesLaquinimod EAE 1 Vez DíapreedtooneNo ratings yet
- 2010 ER Stress Induction in T CellDocument6 pages2010 ER Stress Induction in T CellRaul ReyesNo ratings yet
- Journal of Biotechnology LectureDocument5 pagesJournal of Biotechnology LectureBobbyGunarsoNo ratings yet
- Spittler Et Al. - 1995 - Influence of Glutamine On The Phenotype and FunctiDocument6 pagesSpittler Et Al. - 1995 - Influence of Glutamine On The Phenotype and FunctiMATHILDE MAGRONo ratings yet
- Articulo 1 InmunogenomicaDocument6 pagesArticulo 1 InmunogenomicaMichelle PoulainNo ratings yet
- Molecular and Electrophysiological Characterization of GFP-Expressing CA1 Interneurons in GAD65-GFP MiceDocument11 pagesMolecular and Electrophysiological Characterization of GFP-Expressing CA1 Interneurons in GAD65-GFP MiceFrontiersNo ratings yet
- Toxoplasmose Latente EsplenomegaliaDocument11 pagesToxoplasmose Latente EsplenomegaliaWeverson LinharesNo ratings yet
- Paper 2017Document6 pagesPaper 2017Kokky KoolNo ratings yet
- Regulatory T Cells in Mouse Periapical LesionsDocument5 pagesRegulatory T Cells in Mouse Periapical LesionsEnrique DíazNo ratings yet
- 1 s2.0 0093691X9090575E MainDocument7 pages1 s2.0 0093691X9090575E MainPâmela FreitasNo ratings yet
- Interferon-Like Virus-Inhibitor Induced in Human Leukocytes by PhytohemagglutininDocument2 pagesInterferon-Like Virus-Inhibitor Induced in Human Leukocytes by Phytohemagglutininhoangphuong08101992No ratings yet
- 2572063Document8 pages2572063Nithin RNo ratings yet
- Light-Induced Photon Emission by Mammalian CellsDocument5 pagesLight-Induced Photon Emission by Mammalian CellsAntonis TzambazakisNo ratings yet
- Doksisiklin SpektrumDocument4 pagesDoksisiklin SpektrumTRI SETYA NINGSIHNo ratings yet
- Genome Characterization of - Positive Escherichia Coli Isolated From Pigs With Postweaning Diarrhea in ChinaDocument11 pagesGenome Characterization of - Positive Escherichia Coli Isolated From Pigs With Postweaning Diarrhea in ChinaSusana AsenciosNo ratings yet
- Science 7518614Document3 pagesScience 7518614何燕No ratings yet
- Crespo2011 PDFDocument6 pagesCrespo2011 PDFDANIELA MALAGÓN MONTAÑONo ratings yet
- Production of Transgenic Goats Expressing Human Coagulation Factor IX in The Mammary Glands After Nuclear Transfer Using Transfected Fetal Fibroblast CellsDocument9 pagesProduction of Transgenic Goats Expressing Human Coagulation Factor IX in The Mammary Glands After Nuclear Transfer Using Transfected Fetal Fibroblast CellsMaira QuintanaNo ratings yet
- Ancylostoma Caninum MTP-1, An Astacin-Like Metalloprotease Secreted by Infective Hookworm Larvae, Is Involved in Tissue MigrationDocument7 pagesAncylostoma Caninum MTP-1, An Astacin-Like Metalloprotease Secreted by Infective Hookworm Larvae, Is Involved in Tissue Migrationbmcn54qdvcNo ratings yet
- Co-signal Molecules in T Cell Activation: Immune Regulation in Health and DiseaseFrom EverandCo-signal Molecules in T Cell Activation: Immune Regulation in Health and DiseaseMiyuki AzumaNo ratings yet
- 235 GertDocument11 pages235 GertcarlitoseverectNo ratings yet
- Immunology00256 0071Document10 pagesImmunology00256 0071carlitoseverectNo ratings yet
- Impaired Cutaneous Immune Responses in Thy-1-De Cient Mice: Related ContentDocument8 pagesImpaired Cutaneous Immune Responses in Thy-1-De Cient Mice: Related ContentcarlitoseverectNo ratings yet
- Satellite Parts Catalog SuitesDocument10 pagesSatellite Parts Catalog SuitescarlitoseverectNo ratings yet
- How An Artist Makes A Career Under Persistent Allegations of Sexual Assault and Rape - NRCDocument15 pagesHow An Artist Makes A Career Under Persistent Allegations of Sexual Assault and Rape - NRCcarlitoseverectNo ratings yet
- 2.0 17-02-IM-f (Installation Manual)Document90 pages2.0 17-02-IM-f (Installation Manual)ARMANDOROSAS100% (1)
- Azeem, Et Al ('21) - Tea Leaves Biochar As A Carrier of Bacillus Cereus Improves The Soil Function And.. (ASE)Document13 pagesAzeem, Et Al ('21) - Tea Leaves Biochar As A Carrier of Bacillus Cereus Improves The Soil Function And.. (ASE)aida-69No ratings yet
- Enfield Saheli: Funding NEWSDocument5 pagesEnfield Saheli: Funding NEWSenfieldclubhouseNo ratings yet
- Certificate of Compliance CSA For Fluke 113 - 114 - 115-116-117Document2 pagesCertificate of Compliance CSA For Fluke 113 - 114 - 115-116-117CHARLES BARRAZA100% (1)
- 2020 Proposal Online CarolingDocument2 pages2020 Proposal Online CarolingPel Vincent CruzNo ratings yet
- ASIYADocument3 pagesASIYASOFIKUL HUUSAINNo ratings yet
- BilcoDocument16 pagesBilcoruloNo ratings yet
- Naphthenic Acid / Naphthenate Stabilized Emulsions and The Influence of Crude Oil ComponentsDocument1 pageNaphthenic Acid / Naphthenate Stabilized Emulsions and The Influence of Crude Oil ComponentsLuis Alberto ChirinosNo ratings yet
- Effective Assessment of Crude Oil Sourness For The MaterialDocument7 pagesEffective Assessment of Crude Oil Sourness For The MaterialRenanLacanaNo ratings yet
- Gordon's Functional HealthDocument2 pagesGordon's Functional HealthReina ArabelleNo ratings yet
- SDM-v1 0Document32 pagesSDM-v1 0Franz NussmannNo ratings yet
- Hotel Majestic Kuala Lumpur Press ReleaseDocument3 pagesHotel Majestic Kuala Lumpur Press Releasesam07rocksNo ratings yet
- Biology Related DocumentDocument16 pagesBiology Related DocumentYunjin AnNo ratings yet
- Egypt Agribusiness Report - Q1 2021Document78 pagesEgypt Agribusiness Report - Q1 2021Mina GeorgeNo ratings yet
- DRRR VocabularyDocument2 pagesDRRR VocabularygailNo ratings yet
- Sore ThroatDocument10 pagesSore Throatapi-551073862No ratings yet
- Heart DiseaseDocument88 pagesHeart DiseaseBruce Lawrence MadeirosNo ratings yet
- 2 2 Bioaivers ExperienceDocument80 pages2 2 Bioaivers Experienceblashyrkh_79No ratings yet
- Soil Moisture Monitoring - One-SheeterDocument2 pagesSoil Moisture Monitoring - One-SheeterSorin BurleaNo ratings yet
- Science Journalism ReviewerDocument15 pagesScience Journalism ReviewerHiyami ChanNo ratings yet
- Hemodialysis Patients' Perceptions of Home Hemodialysis and Self-CareDocument13 pagesHemodialysis Patients' Perceptions of Home Hemodialysis and Self-CareRandee BoiiNo ratings yet
- Ethics in Healthcare Setting (Therapy)Document10 pagesEthics in Healthcare Setting (Therapy)Milcah ShabanjiNo ratings yet
- DRRR2324 Q4 DRRDocument7 pagesDRRR2324 Q4 DRRh4gtg7n55gNo ratings yet
- Andovercontinuumul 864 Bacnetsmokecontrolsystemdesignguide 101134Document124 pagesAndovercontinuumul 864 Bacnetsmokecontrolsystemdesignguide 101134Chong CongNo ratings yet
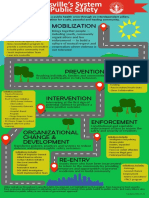
- Reimagining Public SafetyDocument1 pageReimagining Public SafetyWAVE 3 NewsNo ratings yet
- Reflection Paper On Inorganic ChemistryDocument4 pagesReflection Paper On Inorganic ChemistryRachelle Joy VerzonNo ratings yet
- BM 4 - Bricks & BlocksDocument30 pagesBM 4 - Bricks & BlocksArch Reem AlzyoudNo ratings yet
- RDSO Controlled Items Transferred To Other Production Units & COREDocument5 pagesRDSO Controlled Items Transferred To Other Production Units & CORERakesh RoshanNo ratings yet