Download as pdf or txt
You might also like
- Test Bank For Quick and Easy Medical Terminology 9th Edition by LeonardDocument15 pagesTest Bank For Quick and Easy Medical Terminology 9th Edition by Leonardamyrileyixgnaoqjmt100% (30)
- Human Physiology From Cells To Systems 9th Edition Sherwood Solutions ManualDocument12 pagesHuman Physiology From Cells To Systems 9th Edition Sherwood Solutions ManualSamanthaStuartijgt100% (44)
- Abdominal Pain Content PDFDocument71 pagesAbdominal Pain Content PDFyanNo ratings yet
- Ns1 Assignment 1 Neuroembryology and NeurohistologyDocument4 pagesNs1 Assignment 1 Neuroembryology and NeurohistologyMark AbrazaldoNo ratings yet
- The Cells of The Nervous SystemDocument14 pagesThe Cells of The Nervous SystemperezmariaferrieNo ratings yet
- Special HistologyDocument64 pagesSpecial HistologyElijah KamaniNo ratings yet
- Dissection 2Document12 pagesDissection 2hadiqaasif01No ratings yet
- Rullida - EdullantesDocument36 pagesRullida - EdullantesJaleah Gwyneth Fernandez EdullantesNo ratings yet
- Rullida - EdullantesDocument36 pagesRullida - EdullantesJaleah Gwyneth Fernandez EdullantesNo ratings yet
- CNS Histology 1Document60 pagesCNS Histology 1naderipourkianaNo ratings yet
- International Standards For Neurological and Functional Classification of Spinal Cord InjuryDocument14 pagesInternational Standards For Neurological and Functional Classification of Spinal Cord InjuryVijay IyerNo ratings yet
- Modern Review: Gross Anatomy of BrainDocument32 pagesModern Review: Gross Anatomy of BrainVENU B ANo ratings yet
- Neuroanatomy, Brainstem - StatPearls - NCBI BookshelfDocument13 pagesNeuroanatomy, Brainstem - StatPearls - NCBI BookshelfKritika MalhotraNo ratings yet
- 02 - CNS DevelopmentDocument3 pages02 - CNS DevelopmentCemre KuzeyNo ratings yet
- Physiological:biological Psychology NotesDocument36 pagesPhysiological:biological Psychology NotesjeynNo ratings yet
- 1.1.6 NeuroanatomyDocument10 pages1.1.6 NeuroanatomyZayan SyedNo ratings yet
- Chapter IIDocument29 pagesChapter IIKurnia pralisaNo ratings yet
- Introduction and Spinal CordDocument27 pagesIntroduction and Spinal Cordapi-196413370% (1)
- Neuroanatomy: NeurogliaDocument4 pagesNeuroanatomy: Neurogliasamiran dekaNo ratings yet
- Neuron Structural UnitDocument113 pagesNeuron Structural Unitathena villaNo ratings yet
- Animal Physiology Organization of The Vertebrate Nervous System Animal BehaviorDocument16 pagesAnimal Physiology Organization of The Vertebrate Nervous System Animal BehaviorAlyza MayoNo ratings yet
- Histo 14 Nervous Tissue 1Document7 pagesHisto 14 Nervous Tissue 1Hashim GhazoNo ratings yet
- Vertebrate Axis FormationDocument4 pagesVertebrate Axis Formationaaliya saaheenNo ratings yet
- Nervous System PDFDocument9 pagesNervous System PDFAfif Al BaalbakiNo ratings yet
- Plasticity and Evolution of Human BrainDocument39 pagesPlasticity and Evolution of Human BrainPhilipine Gamer And moreNo ratings yet
- Neuroanatomy: Morphology of The BrainDocument19 pagesNeuroanatomy: Morphology of The BrainAsad_Siddiqi_1076No ratings yet
- Class NotesDocument14 pagesClass NotesKovács EnikõNo ratings yet
- Review Article TekuDocument9 pagesReview Article Tekushubhamatilkar04No ratings yet
- Table of Content: Seria L No Page NoDocument48 pagesTable of Content: Seria L No Page NoHadia NoorNo ratings yet
- Endocrine PP NoteDocument9 pagesEndocrine PP NotessinpaibulNo ratings yet
- Evolution of NeurogliaDocument32 pagesEvolution of NeurogliaMichael Rosales VilcaNo ratings yet
- Fundamentals of Nervous System Conduction: B. Pathophysiology 1.) Anatomy and PhysiologyDocument27 pagesFundamentals of Nervous System Conduction: B. Pathophysiology 1.) Anatomy and PhysiologyJanine P. Dela CruzNo ratings yet
- Physiological Psy LectureDocument125 pagesPhysiological Psy LectureKdrama SweetieNo ratings yet
- Neuron and Glial CellsDocument8 pagesNeuron and Glial CellsMurilo Bonet100% (1)
- Catatan Dokter Muda Anatomy OtakDocument23 pagesCatatan Dokter Muda Anatomy OtakFerdi StefiyanNo ratings yet
- Chapter 1 Nerve Cells and Nerve Impulses 2Document19 pagesChapter 1 Nerve Cells and Nerve Impulses 2REPALDA, Irish N.No ratings yet
- Second Quarter Grade 10 - Science HANDOUT No.2 On Neurons, Reflex, Reflex Arc & Brain General Information About The Nervous SystemDocument2 pagesSecond Quarter Grade 10 - Science HANDOUT No.2 On Neurons, Reflex, Reflex Arc & Brain General Information About The Nervous SystemMartin ReaporNo ratings yet
- Brain Mind and - ConsciousnessDocument35 pagesBrain Mind and - ConsciousnessMaxamed Yuusuf DhagjarNo ratings yet
- Physiological Psychology: Physiological Psychology Studies TheDocument18 pagesPhysiological Psychology: Physiological Psychology Studies The31316969No ratings yet
- NeuroscienceDocument169 pagesNeuroscienceSalam Matalka100% (18)
- Cerebellum TextDocument7 pagesCerebellum TextJean Pierre Chastre LuzaNo ratings yet
- Chapter 2 UNIT 1Document19 pagesChapter 2 UNIT 1Ayen LatosaNo ratings yet
- Cnsbloodsupply NotesDocument4 pagesCnsbloodsupply Notescare3ashNo ratings yet
- Trends in Neural EvolutionDocument35 pagesTrends in Neural EvolutionTejpal Dahiya100% (1)
- Histology - Nerve Tissue and The Nervous SystemDocument21 pagesHistology - Nerve Tissue and The Nervous SystemDan UvarovNo ratings yet
- A Brief Introduction To Basic FunctionalDocument50 pagesA Brief Introduction To Basic FunctionalkiflomNo ratings yet
- Animal Nervous System: Logo HereDocument32 pagesAnimal Nervous System: Logo HereAnne Claudette Capin TeofiloNo ratings yet
- Nerve-Part 1Document30 pagesNerve-Part 1GiftNo ratings yet
- Topic 02A - The NeuronsDocument5 pagesTopic 02A - The NeuronsjmvengineerconsNo ratings yet
- Psy 413 ProjectDocument38 pagesPsy 413 ProjectDumebi AneneNo ratings yet
- Physio. Psych Structure-Of-The-Nervous-System-Fact-SheetDocument7 pagesPhysio. Psych Structure-Of-The-Nervous-System-Fact-SheetJustynmae JusainNo ratings yet
- General Appearance ThalamusDocument7 pagesGeneral Appearance ThalamusYubi KimNo ratings yet
- SGTCN ch2Document8 pagesSGTCN ch2Francisco MtzNo ratings yet
- REVIEWER - Biological PsychologyDocument59 pagesREVIEWER - Biological PsychologyMendoza Bernalyn DCNo ratings yet
- Nervous System: Khaleel Alyahya, PHD, Med King Saud University School of Medicine @khaleelyaDocument31 pagesNervous System: Khaleel Alyahya, PHD, Med King Saud University School of Medicine @khaleelyaSalem AlshahraniNo ratings yet
- Three Main Functions:: The Nervous System Is The Master Controlling System of The BodyDocument34 pagesThree Main Functions:: The Nervous System Is The Master Controlling System of The Bodyapi-19641337No ratings yet
- Nervous Tissue LectureDocument55 pagesNervous Tissue Lecturedoubleyouem2003No ratings yet
- THE Cerebellum : Rabbia YousafDocument24 pagesTHE Cerebellum : Rabbia YousafShafaqat Ghani Shafaqat GhaniNo ratings yet
- Anatomy of Nervous TissueDocument54 pagesAnatomy of Nervous TissueNand PrakashNo ratings yet
- Evidence-Based NursingDocument9 pagesEvidence-Based NursingKate AbadNo ratings yet
- The Radiology Assistant - Basic InterpretationDocument67 pagesThe Radiology Assistant - Basic InterpretationMaria LimaNo ratings yet
- 实验项目名称 F ac tors Affecting Urinary Formation: 1. Aim: 2. Subject: Rabbit 3. PrincipleDocument6 pages实验项目名称 F ac tors Affecting Urinary Formation: 1. Aim: 2. Subject: Rabbit 3. PrincipleWai Kwong ChiuNo ratings yet
- Articulo 4Document8 pagesArticulo 4Sonia GarcíaNo ratings yet
- Nervous SystemDocument79 pagesNervous SystemAffan ElahiNo ratings yet
- BRAIN ASYMETRY Exp Rev ProtDocument11 pagesBRAIN ASYMETRY Exp Rev ProtChristina MountakiNo ratings yet
- STS - Stem Cell TherapyDocument29 pagesSTS - Stem Cell TherapyMec AmilasanNo ratings yet
- Casos Clinicos NEJM 2018Document287 pagesCasos Clinicos NEJM 2018Eduarda RosasNo ratings yet
- PathoPHYSIOLOGY OF Spasticity PDFDocument9 pagesPathoPHYSIOLOGY OF Spasticity PDFmandeep axonNo ratings yet
- Frey Syndrome - StatPearlsDocument12 pagesFrey Syndrome - StatPearlsscribdarnabNo ratings yet
- Gladys Cheing RS3030 CNN Introduction To Neuroscience Note 2021Document73 pagesGladys Cheing RS3030 CNN Introduction To Neuroscience Note 2021Tsang AmyNo ratings yet
- Renal Cortical CystDocument2 pagesRenal Cortical Cystra222j239No ratings yet
- Trauma EssayDocument18 pagesTrauma Essaysamakashif1No ratings yet
- The Neuro Ophthalmology Survival Guide Second Edition Edition Burdon Ebook Full ChapterDocument51 pagesThe Neuro Ophthalmology Survival Guide Second Edition Edition Burdon Ebook Full Chapterjuan.small935100% (20)
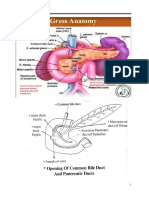
- Carcinoma of PancreasDocument15 pagesCarcinoma of Pancreasعمار عارفNo ratings yet
- Keys To Brain Body BalanceDocument189 pagesKeys To Brain Body BalanceSandeepGambhir100% (1)
- Cortisol, Fixing ImbalancesDocument21 pagesCortisol, Fixing ImbalancesTheresa Dale100% (2)
- Module 1.1 - Introduction To Biological Psychology and The Nerve CellsDocument8 pagesModule 1.1 - Introduction To Biological Psychology and The Nerve CellsJoselitz Reyes TumulakNo ratings yet
- Autopsy Report For Raymond MattiaDocument12 pagesAutopsy Report For Raymond MattiaMichael CooperNo ratings yet
- Atlas of Spleen Pathology and Histhology CompletDocument173 pagesAtlas of Spleen Pathology and Histhology CompletAncamed NycoletaNo ratings yet
- Brain Tumors غﺎﻣﺪﻟا ماروا Definition Brain Tumor ﻒﯾﺮﻌﺘﻟاDocument7 pagesBrain Tumors غﺎﻣﺪﻟا ماروا Definition Brain Tumor ﻒﯾﺮﻌﺘﻟامحمد سعد طه احمدNo ratings yet
- Retinal Vasculitis DDDocument17 pagesRetinal Vasculitis DDPopuri SandhyaNo ratings yet
- Research Secondary HypertensionDocument51 pagesResearch Secondary HypertensionDomeng TalksNo ratings yet
- Kadar Fibrinogen Sebagai Faktor Prognostik Pasien Perdarahan Intraserebral SpontanDocument5 pagesKadar Fibrinogen Sebagai Faktor Prognostik Pasien Perdarahan Intraserebral Spontanannisa edwarNo ratings yet
- 2021 - Uddin - Cognitive and Behavioural Flexibility - Neural Mechanisms and Clinical ConsiderationsDocument13 pages2021 - Uddin - Cognitive and Behavioural Flexibility - Neural Mechanisms and Clinical ConsiderationsjesicaNo ratings yet
- Applied Anatomy of Upper Limb Part OneDocument66 pagesApplied Anatomy of Upper Limb Part Oneprinceej100% (3)
- Endoscopy and ColonosDocument17 pagesEndoscopy and ColonosKim RamosNo ratings yet
- Chapter 6. HistologyDocument22 pagesChapter 6. Histologymaryelle conejarNo ratings yet
- Complication of Neck Dissection & Its Management.: Dr. Sanjay Maharjan 1 Yr Resident, Ent-Hns, MTH, PokharaDocument37 pagesComplication of Neck Dissection & Its Management.: Dr. Sanjay Maharjan 1 Yr Resident, Ent-Hns, MTH, PokharaNitin SharmaNo ratings yet