Download as pdf or txt
You might also like
- Modern Blood Banking & Transfusion Practices 7th Ed PDFDocument699 pagesModern Blood Banking & Transfusion Practices 7th Ed PDFglenn0dela0cruz96% (46)
- Kathleen Langreo Notes FB (Feb6)Document27 pagesKathleen Langreo Notes FB (Feb6)Kaycee Ayo100% (4)
- ALK-positive Lung Cancer: A Moving TargetDocument16 pagesALK-positive Lung Cancer: A Moving TargetcerbulcarpatinNo ratings yet
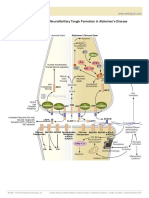
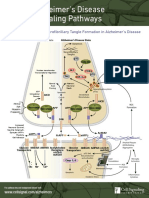
- Amyloid Plaque and Neurofibrillary Tangle Formation in Alzheimer's DiseaseDocument2 pagesAmyloid Plaque and Neurofibrillary Tangle Formation in Alzheimer's DiseaseMelanieNo ratings yet
- Vasodilation: Vasodilation Is The Widening of Blood VesselsDocument8 pagesVasodilation: Vasodilation Is The Widening of Blood VesselsNTA UGC-NETNo ratings yet
- NOTZ SUMMARY - Aldosterone TableDocument1 pageNOTZ SUMMARY - Aldosterone TableAdmin DutiesNo ratings yet
- Overview - Regulation of ApoptosisDocument1 pageOverview - Regulation of ApoptosisAym mantezNo ratings yet
- Structural and Molecular Bases of Cardiac Inward Rectifier Potassium Channel FunctionDocument11 pagesStructural and Molecular Bases of Cardiac Inward Rectifier Potassium Channel FunctionArmando RodriguezNo ratings yet
- Https - Learn - Cellsignal.com - Hubfs - Pdfs - 15 Fly Alzh Ref m096 Eng 00 Alzheimers PW Handout DigitalDocument2 pagesHttps - Learn - Cellsignal.com - Hubfs - Pdfs - 15 Fly Alzh Ref m096 Eng 00 Alzheimers PW Handout DigitaltamaraNo ratings yet
- Table 4-2. Mechanism of Action of Selected Nonpeptide NeurotransmittersDocument1 pageTable 4-2. Mechanism of Action of Selected Nonpeptide NeurotransmittersaustinchenNo ratings yet
- Extrinsic Cardiac Conduction System AtfDocument2 pagesExtrinsic Cardiac Conduction System AtfMariaNo ratings yet
- Cardiovascular - Electrophysiology - Extrinsic Cardiac Conduction SystemDocument2 pagesCardiovascular - Electrophysiology - Extrinsic Cardiac Conduction SystemSaransh JainNo ratings yet
- FCPS Surgery 25 Aug 2023 (M)Document20 pagesFCPS Surgery 25 Aug 2023 (M)Tauseef AfridiNo ratings yet
- Ncomms2283 Página 2Document1 pageNcomms2283 Página 2OscarNo ratings yet
- Calcium For Resuscitation?: E. Erdmann and E. Reuschel-JanetschekDocument7 pagesCalcium For Resuscitation?: E. Erdmann and E. Reuschel-JanetschekMislav HilcNo ratings yet
- 1 s2.0 S0005273621000687 MainDocument7 pages1 s2.0 S0005273621000687 MainMarco AsprónNo ratings yet
- Transmitter PropertiesDocument1 pageTransmitter PropertiesNaveed PashaNo ratings yet
- Frischauf2008 STIM Orai CouplingDocument8 pagesFrischauf2008 STIM Orai CouplingGiulia AndreeaNo ratings yet
- NT Release PPCNS 2023Document16 pagesNT Release PPCNS 2023Yasmin Sophia MNo ratings yet
- CC2 TransDocument12 pagesCC2 TransAnathalea ReyesNo ratings yet
- Colledge Trnds Cell Biol 1999 p216Document6 pagesColledge Trnds Cell Biol 1999 p216KRIESHA 198No ratings yet
- 1646 FullDocument16 pages1646 FullMedranoReyesLuisinNo ratings yet
- Molecular Physiology of Cardiac Repolarization: Nerbonne, Jeanne M., and Robert S. KassDocument49 pagesMolecular Physiology of Cardiac Repolarization: Nerbonne, Jeanne M., and Robert S. Kassrizki sundusiasihNo ratings yet
- Molluscan: Inhibition of Protein Synthesis Prolongs Ca2 - Mediated Reduction of K+ Currents in NeuronsDocument5 pagesMolluscan: Inhibition of Protein Synthesis Prolongs Ca2 - Mediated Reduction of K+ Currents in NeuronsAmale RachaNo ratings yet
- Activation of PKA and Epac Proteins by Cyclic AMP Depletes Intracellular Calcium Stores and Reduces Calcium Availability For VasoconstrictionDocument30 pagesActivation of PKA and Epac Proteins by Cyclic AMP Depletes Intracellular Calcium Stores and Reduces Calcium Availability For VasoconstrictionLeonel LedezmaNo ratings yet
- Exercise and Fatigue: Integrating The Role of K, Na and CL in The Regulation of Sarcolemmal Excitability of Skeletal MuscleDocument34 pagesExercise and Fatigue: Integrating The Role of K, Na and CL in The Regulation of Sarcolemmal Excitability of Skeletal MuscleAlejandroNo ratings yet
- Nitric Oxide SignalingDocument1 pageNitric Oxide SignalingAlaa HassanNo ratings yet
- Https Learn - Cellsignal.com Hubfs Pdfs 19-Cel-11131-Pwh-DigitalDocument2 pagesHttps Learn - Cellsignal.com Hubfs Pdfs 19-Cel-11131-Pwh-DigitaltamaraNo ratings yet
- MMC 2Document30 pagesMMC 2ydv.deepika11297No ratings yet
- Cell DeathDocument13 pagesCell DeathTarunSreerajNo ratings yet
- Https - Learn - Cellsignal.com - Hubfs - Pdfs - 16 Fly DBL 00063 Eng WNT B Catenin Diff PW Handout SMDocument2 pagesHttps - Learn - Cellsignal.com - Hubfs - Pdfs - 16 Fly DBL 00063 Eng WNT B Catenin Diff PW Handout SMtamaraNo ratings yet
- Central Synapse and Neuromuscular Junction: Same Players, Different RolesDocument8 pagesCentral Synapse and Neuromuscular Junction: Same Players, Different RolesMarcelo Pires de OliveiraNo ratings yet
- Synapse Neurobiology: Week 6Document37 pagesSynapse Neurobiology: Week 6bobobNo ratings yet
- M - PH: The Relationship Between Length and Active Force of A Skeletal MuscleDocument11 pagesM - PH: The Relationship Between Length and Active Force of A Skeletal MuscleFlowerNo ratings yet
- Creatine Kinase: NAC. Kinetic UV. LiquidDocument2 pagesCreatine Kinase: NAC. Kinetic UV. LiquidJorge PinedaNo ratings yet
- OS214 20060306 Grp10a Chronic Renal FailureDocument3 pagesOS214 20060306 Grp10a Chronic Renal Failureapi-3799593100% (5)
- SKINDocument1 pageSKINHafshary D. ThanialNo ratings yet
- 1 s2.0 0092867495904085 Main - CompressedDocument10 pages1 s2.0 0092867495904085 Main - CompressedBinada EnjelitaNo ratings yet
- Nihms 181336Document6 pagesNihms 181336Giulia AndreeaNo ratings yet
- Abnormal Intrastore Calcium Signaling in Chronic Heart FailureDocument6 pagesAbnormal Intrastore Calcium Signaling in Chronic Heart FailureFrontiersNo ratings yet
- Ahnak, a new player in β-adrenergic regulation of the cardiac L-type Ca channelDocument7 pagesAhnak, a new player in β-adrenergic regulation of the cardiac L-type Ca channeliuventasNo ratings yet
- In The Name of ALLAH, The Beneficent The MercifulDocument139 pagesIn The Name of ALLAH, The Beneficent The Mercifulkhawar93No ratings yet
- 5 - Heart - EcgDocument83 pages5 - Heart - EcgStephen PramatyaNo ratings yet
- HypokalemiaDocument1 pageHypokalemiaArif AlvaNo ratings yet
- Exinator2: Whole Genome Somatic MutationsDocument1 pageExinator2: Whole Genome Somatic MutationsshoummowNo ratings yet
- Cardio Electrophysiology (Preview) PDFDocument11 pagesCardio Electrophysiology (Preview) PDFSuwijak MeenapaNo ratings yet
- Kalium EInfluss Auf HerzDocument1 pageKalium EInfluss Auf HerzAlexandra AlexaNo ratings yet
- Adreasen 2002 Inhibition of Slow Ca Activated K Current 4AP Rat CA1Document13 pagesAdreasen 2002 Inhibition of Slow Ca Activated K Current 4AP Rat CA1JortegloriaNo ratings yet
- Cryo-EM Reveals The Transition of Arp2-3 Complex From Inactive To Nucleation-Competent StateDocument24 pagesCryo-EM Reveals The Transition of Arp2-3 Complex From Inactive To Nucleation-Competent StateKarina B Hernandez ANo ratings yet
- T Cell Receptor Signaling: F-A CtinDocument2 pagesT Cell Receptor Signaling: F-A CtinOlgaNo ratings yet
- Excitation-Contraction Coupling in Vertebrate Skeletal Muscle: A Tale of Two Calcium Channels MinireviewDocument4 pagesExcitation-Contraction Coupling in Vertebrate Skeletal Muscle: A Tale of Two Calcium Channels MinireviewHari SofiaNo ratings yet
- 2017-29-Digoxin and Amiodarone (A Vorster)Document10 pages2017-29-Digoxin and Amiodarone (A Vorster)Sadiq ZakariaNo ratings yet
- Individual AssignmentDocument3 pagesIndividual AssignmentPutri Rizky AmaliaNo ratings yet
- CRISPR-Cas12a Delivery by DNA-mediated Bioresponsive Editing For CholesterolDocument25 pagesCRISPR-Cas12a Delivery by DNA-mediated Bioresponsive Editing For CholesterolTanzeela noureenNo ratings yet
- SARS-CoV-2 Variants - UK + South African + Brazil Variants - Key AtfDocument1 pageSARS-CoV-2 Variants - UK + South African + Brazil Variants - Key AtfJuan Manuel Tapia AlzateNo ratings yet
- KSP44 / KSP45 NPN Epitaxial Silicon Transistor: FeaturesDocument8 pagesKSP44 / KSP45 NPN Epitaxial Silicon Transistor: FeaturesIIIkwarkaNo ratings yet
- Group SelectionDocument12 pagesGroup SelectionNTA UGC-NETNo ratings yet
- The HEAT Repeat Protein HPO-27 Is A Lysosome Fission Factor: ArticleDocument32 pagesThe HEAT Repeat Protein HPO-27 Is A Lysosome Fission Factor: Articleadnanjamal350No ratings yet
- Epistasis Answer KeyDocument5 pagesEpistasis Answer KeyIs noorNo ratings yet
- How To Interpret A KaryotypeDocument2 pagesHow To Interpret A KaryotypeLiberty NguyenNo ratings yet
- Gene TherapyDocument6 pagesGene Therapyjomari palmeroNo ratings yet
- Lesson 1 Characteristics of LifeDocument5 pagesLesson 1 Characteristics of LifeKrish D.No ratings yet
- Year-12-Biology Excel Revision Cards PDFDocument109 pagesYear-12-Biology Excel Revision Cards PDFTylerNo ratings yet
- Science Animal PortfolioDocument9 pagesScience Animal PortfolioCrys TalNo ratings yet
- Self Help (Non-Fiction)Document7 pagesSelf Help (Non-Fiction)jtwendtNo ratings yet
- Earth and Life Science: Quarter 2 - Module 3Document27 pagesEarth and Life Science: Quarter 2 - Module 3Jerald De La Cruz100% (2)
- Discussion of Modernization Li Lu PDFDocument124 pagesDiscussion of Modernization Li Lu PDFanchandra88No ratings yet
- 12 (Agriculture) Set-A 1Document36 pages12 (Agriculture) Set-A 1Himanshu bishtNo ratings yet
- Techno-1.3-The Molecular Basis of HeredityDocument30 pagesTechno-1.3-The Molecular Basis of HeredityMark DeGrootteNo ratings yet
- 2 5 MacroevolutionDocument32 pages2 5 MacroevolutionJOCELYN JOSE IMS22136No ratings yet
- CSE 516/CSE 446 Introduction To Bioinformatics: Presented by Dr. Shazzad Hosain Asst. Prof. EECS, NSUDocument25 pagesCSE 516/CSE 446 Introduction To Bioinformatics: Presented by Dr. Shazzad Hosain Asst. Prof. EECS, NSUAlimushwan AdnanNo ratings yet
- Genetics NotesDocument20 pagesGenetics Noteslebohangcandy8No ratings yet
- A Report On The Mosquitoes of Mainland Åland, Southwestern Finland and Revised List of Finnish Mosquitoes - Culverwell C.Document10 pagesA Report On The Mosquitoes of Mainland Åland, Southwestern Finland and Revised List of Finnish Mosquitoes - Culverwell C.Ece Zeynep AkdemirNo ratings yet
- Hall, Young, Kenway - 2002 - Manual For The Determination of Egg Fertility in Penaeus Monodon - PDF TraducidoDocument54 pagesHall, Young, Kenway - 2002 - Manual For The Determination of Egg Fertility in Penaeus Monodon - PDF Traducidoelina del mar rodriguez barbosaNo ratings yet
- Tesis Doctoral - Iván Carreño RuizDocument237 pagesTesis Doctoral - Iván Carreño RuizJaime AcostaNo ratings yet
- Biotechnology Applications: by Professor Tareq Alasadi Alkut University CollegeDocument7 pagesBiotechnology Applications: by Professor Tareq Alasadi Alkut University CollegePrama Hartami100% (1)
- New - Mechanisms of SpeciationDocument33 pagesNew - Mechanisms of SpeciationAidyn TemirbekovaNo ratings yet
- ViewpdfDocument11 pagesViewpdfBhanu Pratap Singh TomarNo ratings yet
- Chromosomes, Genes and DnaDocument3 pagesChromosomes, Genes and DnaRaahilNo ratings yet
- Graduate Aptitude Test-Biotechnology (Gat-B) 2021 Parts / Sections Number of Questions Number of Questions To Be Attempted MarksDocument2 pagesGraduate Aptitude Test-Biotechnology (Gat-B) 2021 Parts / Sections Number of Questions Number of Questions To Be Attempted MarksSamarth SinghNo ratings yet
- General Biology 2: Quarter 3, Module 1 Genetic EngineeringDocument20 pagesGeneral Biology 2: Quarter 3, Module 1 Genetic EngineeringRonalyn AndaganNo ratings yet
- Ga 19Document15 pagesGa 19api-676767477No ratings yet
- BIO345 Syllabus Fall 2023-Byblos CampusDocument7 pagesBIO345 Syllabus Fall 2023-Byblos Campusgaelle tannousNo ratings yet
- Activity No.1 BSci 101Document3 pagesActivity No.1 BSci 101Shamaica SurigaoNo ratings yet
- Biology-WPS OfficeDocument2 pagesBiology-WPS OfficeLyca FelicisimoNo ratings yet
- Zoology (3 Marks Questions)Document4 pagesZoology (3 Marks Questions)Barani JanarthanamNo ratings yet