Download as pdf or txt
You might also like
- NS1881 EXAM PREP Flashcards - QuizletDocument3 pagesNS1881 EXAM PREP Flashcards - QuizletLaetitia ReyesNo ratings yet
- PVJ PDFDocument6 pagesPVJ PDFAshwini Kumar JhaNo ratings yet
- Canine Demodecosis and Babesiosis - Bambang Pontjo-INDONESIADocument14 pagesCanine Demodecosis and Babesiosis - Bambang Pontjo-INDONESIAIkhwanul KhairiaNo ratings yet
- (2017) BELOUSOVA, Irena E. Mycosis Fungoides Manifesting As Giant Cell Lichenoid DermatitisDocument3 pages(2017) BELOUSOVA, Irena E. Mycosis Fungoides Manifesting As Giant Cell Lichenoid DermatitisCassandra VérasNo ratings yet
- Hongos PerroDocument4 pagesHongos PerroHelen Dellepiane GilNo ratings yet
- Discoid Lupus Erythematosus (DLE) in A Spitz DogDocument3 pagesDiscoid Lupus Erythematosus (DLE) in A Spitz DogPatrícia StancuNo ratings yet
- Parakeratosis Variegata: A Possible Role of Environmental Hazards?Document4 pagesParakeratosis Variegata: A Possible Role of Environmental Hazards?Ega AihenaNo ratings yet
- Egert2017skinmicrobiota 220829 170221Document8 pagesEgert2017skinmicrobiota 220829 170221Leandro QuitoNo ratings yet
- Feline Plasma Cell Pododermatitis - A Study of 8 Cases (Pages 333-337) PDFDocument5 pagesFeline Plasma Cell Pododermatitis - A Study of 8 Cases (Pages 333-337) PDFjenNo ratings yet
- Trichoscopic Diagnosis of Cutaneous Pelodera Strongyloides Infestation in A DogDocument5 pagesTrichoscopic Diagnosis of Cutaneous Pelodera Strongyloides Infestation in A DogRebeka MenezesNo ratings yet
- Dermatitis Parasitaria en PerrosDocument5 pagesDermatitis Parasitaria en PerrosMauricio Urbina diazNo ratings yet
- Bizkova 2009Document8 pagesBizkova 2009mario256No ratings yet
- Batrachochytrium Dendrobatidis Description 1999Document10 pagesBatrachochytrium Dendrobatidis Description 1999bsdunleyNo ratings yet
- 1385Document7 pages1385Vlad BudaeNo ratings yet
- A Case of Feline Panleukopenia in Felis Silvestris in Iran Confirmed by PCRDocument5 pagesA Case of Feline Panleukopenia in Felis Silvestris in Iran Confirmed by PCRintan noorNo ratings yet
- Chapter 1 - OverviewDocument47 pagesChapter 1 - OverviewZea Mae AgirNo ratings yet
- Case ReportDocument4 pagesCase ReportbejoNo ratings yet
- Morphogenesis of Koi Herpesvirus Observed by Electron MicrosDocument8 pagesMorphogenesis of Koi Herpesvirus Observed by Electron MicrosgiuseppegnrNo ratings yet
- Prepared By: Ken Robin A. Canada, RMT, Mls AscpiDocument35 pagesPrepared By: Ken Robin A. Canada, RMT, Mls AscpiKen Robin CanadaNo ratings yet
- Granuloma Is Defined As A Circumscribed, Tiny Lesion, About 1 MM in Diameter, Composed PredominantlyDocument6 pagesGranuloma Is Defined As A Circumscribed, Tiny Lesion, About 1 MM in Diameter, Composed PredominantlyIsak ShatikaNo ratings yet
- Cutaneous Lesions of The Canine ScrotumDocument14 pagesCutaneous Lesions of The Canine ScrotumLuciana Diegues100% (1)
- Sheep PaperDocument10 pagesSheep Paperbilal1311511382No ratings yet
- Innate Immunity, Microbiology, Inflammation - ABSTRACTSDocument1 pageInnate Immunity, Microbiology, Inflammation - ABSTRACTSFebrian AlfaroNo ratings yet
- Apoptosis in Selected Skin Diseases PDFDocument9 pagesApoptosis in Selected Skin Diseases PDFDiana Gabriela GutuerrezNo ratings yet
- Epidermodysplasia-Verruciformis YmjdDocument1 pageEpidermodysplasia-Verruciformis YmjdAndrés WunderwaldNo ratings yet
- Demodex Mites - Commensals, Parasites or Mutualistic Organisms?Document3 pagesDemodex Mites - Commensals, Parasites or Mutualistic Organisms?SMA Negeri 1 SedayuNo ratings yet
- Pi Is 0190962208009328Document2 pagesPi Is 0190962208009328Nabhya MaheshwariNo ratings yet
- Invasive Epithelial Mesothelioma in A Dog: F. R, B. B, K. R, E. S, D. BDocument5 pagesInvasive Epithelial Mesothelioma in A Dog: F. R, B. B, K. R, E. S, D. BjamesyuNo ratings yet
- Ciclosporin Therapy For Canine Generalized Discoid LupusDocument6 pagesCiclosporin Therapy For Canine Generalized Discoid LupusPatrícia StancuNo ratings yet
- Pediatric Dermatology - 2022 - Li - Lipoid Proteinosis Novel ECM1 Pathogenic Variants and Intrafamilial Variability inDocument7 pagesPediatric Dermatology - 2022 - Li - Lipoid Proteinosis Novel ECM1 Pathogenic Variants and Intrafamilial Variability inArianny SilvaNo ratings yet
- Ref 3 Skin Changes PMDocument5 pagesRef 3 Skin Changes PMfa.anselmo89No ratings yet
- Tsai (2007) - NSDocument2 pagesTsai (2007) - NSIlma ArifaniNo ratings yet
- A Case of Feline Panleukopenia in Felis Silvestris in Iran Confirmed by PCRDocument4 pagesA Case of Feline Panleukopenia in Felis Silvestris in Iran Confirmed by PCRUsamah El-habibNo ratings yet
- BABUDIERI 1972 (Hemoparásitos)Document6 pagesBABUDIERI 1972 (Hemoparásitos)Mariana CabagnaNo ratings yet
- Periradicular Lesions (Chapter 5)Document28 pagesPeriradicular Lesions (Chapter 5)revlieeNo ratings yet
- Buffalo LeprosyDocument4 pagesBuffalo LeprosyALTAF HUSAINNo ratings yet
- Stau Dinger 2011Document9 pagesStau Dinger 2011inforNo ratings yet
- Krok Key WordsDocument50 pagesKrok Key WordsKeller 0No ratings yet
- The Etiology and Treatment of Erythrasma : Mellit - UsDocument8 pagesThe Etiology and Treatment of Erythrasma : Mellit - Usriska_leilyNo ratings yet
- Histopathological Characterization of Induced Paederus Dermatitis Caused by Egyptian Rove Beetles (Paederus Alfierii)Document6 pagesHistopathological Characterization of Induced Paederus Dermatitis Caused by Egyptian Rove Beetles (Paederus Alfierii)rositaNo ratings yet
- Barrier Function, Epidermal Differentiation, and Human B-Defensin 2 Expression in Tinea CorporisDocument8 pagesBarrier Function, Epidermal Differentiation, and Human B-Defensin 2 Expression in Tinea CorporisMuhammad HafizuddinNo ratings yet
- Case Report: Subcorneal Pustular Dermatosis in Childhood: A Case Report and Review of The LiteratureDocument6 pagesCase Report: Subcorneal Pustular Dermatosis in Childhood: A Case Report and Review of The LiteratureIrsalinaHusnaAzwirNo ratings yet
- A Case Report On Scabies in A GoatDocument4 pagesA Case Report On Scabies in A Goatalifnaufaliyanto100% (1)
- Jin 2009 Models of Atopic DermatitisDocument10 pagesJin 2009 Models of Atopic DermatitisBrigitta SzöllősiNo ratings yet
- 10.1007@s40257 017 0309 6Document3 pages10.1007@s40257 017 0309 6shf.mxlikNo ratings yet
- #AW - Cooper, 2003Document13 pages#AW - Cooper, 2003Andi WirdaniNo ratings yet
- Cutaneous GranulomasDocument2 pagesCutaneous GranulomasloreagubenjiNo ratings yet
- Effect of A Velogenic Newcastle Disease Virus On Body and Organ Weights of Vaccinated Shika Brown CocksDocument6 pagesEffect of A Velogenic Newcastle Disease Virus On Body and Organ Weights of Vaccinated Shika Brown CocksCryzta Privit SenorinNo ratings yet
- Cutaneous MycosesDocument36 pagesCutaneous Mycosesrhosie AquinoNo ratings yet
- Histopathological Study of Endometritis of The Cows: AbstractsDocument6 pagesHistopathological Study of Endometritis of The Cows: Abstractsهدير عبدالمطلب حمزةNo ratings yet
- Celule DendriticeDocument9 pagesCelule DendriticeDianaNo ratings yet
- Nonbullous Pemphigoid: Prodrome of Bullous Pemphigoid or A Distinct Pemphigoid Variant?Document7 pagesNonbullous Pemphigoid: Prodrome of Bullous Pemphigoid or A Distinct Pemphigoid Variant?FatimaNo ratings yet
- Experimental Infection ofDocument6 pagesExperimental Infection ofNurrahmah HijrahNo ratings yet
- CoccidosisDocument288 pagesCoccidosisMegersaNo ratings yet
- Leptospirosis A Re-Emerging DiseaseDocument4 pagesLeptospirosis A Re-Emerging DiseasemelshiyafNo ratings yet
- Radiation Tolerance of The SkinDocument21 pagesRadiation Tolerance of The SkinYEREMY ALEXANDER FERNÁNDEZNo ratings yet
- Otomycosis: Yan Edward, Dolly IrfandyDocument6 pagesOtomycosis: Yan Edward, Dolly IrfandyM Reza Kurnia AliNo ratings yet
- UP Devon Rex Vet DermDocument7 pagesUP Devon Rex Vet Dermandrostona980No ratings yet
- Non-inflammatory immunology: An introduction to the immune system and its pathologiesFrom EverandNon-inflammatory immunology: An introduction to the immune system and its pathologiesNo ratings yet
- Triplebottom Hides and Skins Quality Requirements.2Document77 pagesTriplebottom Hides and Skins Quality Requirements.2John WaweruNo ratings yet
- NSO DermabrasionDocument4 pagesNSO DermabrasionIman TawasilNo ratings yet
- POSAS Patient Scale: The Patient and Observer Scar Assessment Scale v2.0 / ENDocument2 pagesPOSAS Patient Scale: The Patient and Observer Scar Assessment Scale v2.0 / ENAde Ewa PermanaNo ratings yet
- Menganalisa Kompetitor Pada Produk MakananDocument18 pagesMenganalisa Kompetitor Pada Produk MakananJeje KediriNo ratings yet
- IGCSE Biology - TOPIC 14 - Endocrine System & Homeostasis SUMMARY NOTESDocument8 pagesIGCSE Biology - TOPIC 14 - Endocrine System & Homeostasis SUMMARY NOTESpeggyyu12345No ratings yet
- Lecture No 1 CRM 223 DactylosDocument25 pagesLecture No 1 CRM 223 Dactylosjanicaritzpaez45No ratings yet
- Gupta Ak Et Al Fungal Skin Infections PDFDocument17 pagesGupta Ak Et Al Fungal Skin Infections PDFValeria BeltranNo ratings yet
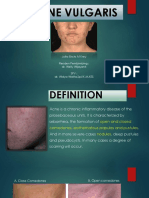
- Acne Vulgaris PowerpointDocument21 pagesAcne Vulgaris PowerpointEricks Manfun100% (3)
- Geria HandoutsDocument43 pagesGeria HandoutsMaria Jerriessa Anne GanadinNo ratings yet
- 176 Anatomy Integumentary SystemDocument29 pages176 Anatomy Integumentary SystemJoana Mae PortezaNo ratings yet
- Basic Science and Sructure of Skin MCQsDocument92 pagesBasic Science and Sructure of Skin MCQsDr.Tawheed88% (17)
- The Integumentary SystemDocument10 pagesThe Integumentary SystemAlliyah GuerreroNo ratings yet
- TSM1501G: Toyota Engineering StandardDocument12 pagesTSM1501G: Toyota Engineering StandardMiguel FelipeNo ratings yet
- Cancer of The Oral CavityDocument1 pageCancer of The Oral CavityAlan James Sy CañizaresNo ratings yet
- 24 Urological Trauma LRDocument67 pages24 Urological Trauma LRManushi HenadeeraNo ratings yet
- Oily Sensitive Skin: A Review of Management OptionsDocument5 pagesOily Sensitive Skin: A Review of Management Optionsthichkhampha_py94No ratings yet
- Homeostasis and ExcretionDocument35 pagesHomeostasis and ExcretionronaldwepukhuluNo ratings yet
- 31 Slide Tap Dich - Session 31Document50 pages31 Slide Tap Dich - Session 31Đào Huỳnh PhúcNo ratings yet
- Newer Understanding of Specific Anatomic Targets in The Aging Face As Applied To Injectables - Aging Changes in The Craniofacial Skeleton and Facial LigamentsDocument5 pagesNewer Understanding of Specific Anatomic Targets in The Aging Face As Applied To Injectables - Aging Changes in The Craniofacial Skeleton and Facial LigamentsArcelino FariasNo ratings yet
- External Parasites of Small RuminantsDocument256 pagesExternal Parasites of Small RuminantsRBlitzoneNo ratings yet
- Nummular EczemaDocument1 pageNummular EczemaCitra AnandiraNo ratings yet
- Functions of The SkinDocument6 pagesFunctions of The SkinMieshka EzekielNo ratings yet
- Admin,+101+ +839+ +Anak+Agung+Indah+Jayanthi+ +galleyDocument10 pagesAdmin,+101+ +839+ +Anak+Agung+Indah+Jayanthi+ +galleySilvia OktavianiNo ratings yet
- DermatoscopieDocument220 pagesDermatoscopiepodacclaudiu100% (2)
- Fischer Et Al-2008-Journal of Pineal ResearchDocument15 pagesFischer Et Al-2008-Journal of Pineal ResearchgeoaislaNo ratings yet
- Buzz Word Cheat Sheet (Derm)Document5 pagesBuzz Word Cheat Sheet (Derm)emilyjvaldez99No ratings yet
- PDFDocument13 pagesPDFUlfah Uph UphNo ratings yet
- Picore Product Introduction (2021) - NewDocument23 pagesPicore Product Introduction (2021) - Newsamuel lumbaNo ratings yet
- Tsubo Vital Points For Oriental Therapy Serizawa KatsusukeDocument251 pagesTsubo Vital Points For Oriental Therapy Serizawa KatsusukeMihail Negoita100% (1)