
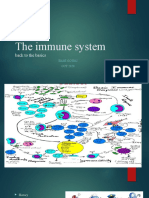
Immune GM Eng 2022
Immune GM Eng 2022
You might also like
- Robbins Pathology - Chapter 3 TransDocument18 pagesRobbins Pathology - Chapter 3 Transnath nath100% (7)
- Chapter 6 - Diseases of The Immune SystemDocument3 pagesChapter 6 - Diseases of The Immune SystemArun Kumar SinghNo ratings yet
- Basic Immunology 2Document66 pagesBasic Immunology 2JabbarTapiheruNo ratings yet
- Tugas Imunologi Dasar Fast TrackDocument23 pagesTugas Imunologi Dasar Fast TrackAnditri WeningtyasNo ratings yet
- Prologue Syllabus 2008Document14 pagesPrologue Syllabus 2008Francisco Eriberto de LimaNo ratings yet
- Lecture 1 - The Immune SystemDocument29 pagesLecture 1 - The Immune SystemGana KhaledNo ratings yet
- Lecturer One Introduction To General Principles of ImmunologyDocument6 pagesLecturer One Introduction To General Principles of ImmunologyKevin KipropNo ratings yet
- 53Document29 pages53Keerthy Venthen100% (1)
- 53 PDFDocument29 pages53 PDFBrunoNo ratings yet
- 1.notes PowerpointDocument27 pages1.notes Powerpointt4ths999No ratings yet
- Basic Immunology From The Dermatologic Point of ViewDocument87 pagesBasic Immunology From The Dermatologic Point of ViewnovelraaaNo ratings yet
- Openstax - Biology2e - ch42 (VVL)Document47 pagesOpenstax - Biology2e - ch42 (VVL)ANo ratings yet
- Immune System 2018Document3 pagesImmune System 2018Luna SartoriNo ratings yet
- Chapter 6 - Diseases of The Immune SystemDocument3 pagesChapter 6 - Diseases of The Immune SystemTurinawe Bin ByensiNo ratings yet
- The Immune System: Back To The BasicsDocument37 pagesThe Immune System: Back To The BasicsHani GowaiNo ratings yet
- BCH444 PDFDocument232 pagesBCH444 PDFYaseen Ahmad100% (1)
- Topic 1,2,3 y 4Document15 pagesTopic 1,2,3 y 4Olaya De Dios HuertaNo ratings yet
- Tugas ImunilogiDocument3 pagesTugas Imunilogilapalo46No ratings yet
- Handout For Innate 1, Innate 2 and AdaptiveDocument26 pagesHandout For Innate 1, Innate 2 and Adaptive7231598No ratings yet
- Prelims - GMJ Lecture - Module 2 III DisorderDocument5 pagesPrelims - GMJ Lecture - Module 2 III DisorderjuiceNo ratings yet
- Study Guide For Introduction To Immune SystemDocument9 pagesStudy Guide For Introduction To Immune SystemOPERAñA ELLAYZA RB DECANONo ratings yet
- Immune System To Agent InfectionDocument70 pagesImmune System To Agent Infectionliza yunitaNo ratings yet
- Chapter 09. Host Response To InfectionDocument51 pagesChapter 09. Host Response To InfectionZin LimNo ratings yet
- Innate and Acquired ImmunityDocument17 pagesInnate and Acquired ImmunityPiyush KumarNo ratings yet
- IMMUNOLOGY Flash PointsDocument3 pagesIMMUNOLOGY Flash PointsHassan AhmadNo ratings yet
- The Immune System:: Innate and Acquired ImmunityDocument47 pagesThe Immune System:: Innate and Acquired ImmunityBad BunnyNo ratings yet
- ImmunologyDocument11 pagesImmunologyBiology BảoNo ratings yet
- DAMPs, PAMPs and Alarmins. All We Need To Know About DangerDocument5 pagesDAMPs, PAMPs and Alarmins. All We Need To Know About DangerjesusNo ratings yet
- Basic Immunology (Revision) : DR Z MakatiniDocument51 pagesBasic Immunology (Revision) : DR Z MakatiniEstee van WykNo ratings yet
- Immunophysiology: DR: Omer Eltahir Registrar of PathologyDocument40 pagesImmunophysiology: DR: Omer Eltahir Registrar of PathologySafa TaweelNo ratings yet
- Immunology and Serology SolutionDocument17 pagesImmunology and Serology Solutionzakaria hossainNo ratings yet
- 12 13 2006 Life Science Introduction To Immune 2006Document99 pages12 13 2006 Life Science Introduction To Immune 2006api-3696530No ratings yet
- Defence SystemDocument33 pagesDefence Systemboneheaded9No ratings yet
- Immune Response (Antibody-Antigen Reactions)Document20 pagesImmune Response (Antibody-Antigen Reactions)SimonNo ratings yet
- M2 Lecture 8 Immune ResponseDocument23 pagesM2 Lecture 8 Immune ResponseChing CHANNo ratings yet
- Ih Lec PrelimDocument24 pagesIh Lec PrelimReann SmithNo ratings yet
- NotesDocument2 pagesNotesdiceNo ratings yet
- Disease of Immune System - Siti Mirhalina 2Document25 pagesDisease of Immune System - Siti Mirhalina 2M.Satria PerdanapNo ratings yet
- Immunology Unveiled: A Comprehensive Journey through the Human Immune System: Guardians of the Body: The Unseen Heroes of ImmunityFrom EverandImmunology Unveiled: A Comprehensive Journey through the Human Immune System: Guardians of the Body: The Unseen Heroes of ImmunityNo ratings yet
- Innate ImmunityDocument16 pagesInnate ImmunityikhaNo ratings yet
- Immunity To Infections Lecture NotesDocument40 pagesImmunity To Infections Lecture Notesfredrick damianNo ratings yet
- Immunology SummaryDocument81 pagesImmunology SummaryDiaz ShehanNo ratings yet
- 2 - The Innate ImmunityDocument4 pages2 - The Innate Immunitynomore891No ratings yet
- Inflammation and Inflammatory MediatorsDocument28 pagesInflammation and Inflammatory Mediatorsla.instagramawaNo ratings yet
- Immunology: Combined Class (Mental Health Psichatry, Clinical Anastasia, & Clinical Sciences) Year 2 30/03/2022Document30 pagesImmunology: Combined Class (Mental Health Psichatry, Clinical Anastasia, & Clinical Sciences) Year 2 30/03/2022Derick NKANDU KALUBANo ratings yet
- ImunologiDocument4 pagesImunologiDhiya NabilaNo ratings yet
- Introduction To ImmunologyDocument14 pagesIntroduction To Immunologykaiyeol exoNo ratings yet
- Immunologi Klinik Ama 09102019 PDFDocument101 pagesImmunologi Klinik Ama 09102019 PDFZas VutraNo ratings yet
- Immunometabolism IDocument29 pagesImmunometabolism IMatheus TavaresNo ratings yet
- Chap 16 Part 3 Antigen-Antibody ComplexesDocument6 pagesChap 16 Part 3 Antigen-Antibody ComplexesclrssNo ratings yet
- 4 Immune DisordersDocument11 pages4 Immune DisorderscedonuliNo ratings yet
- General Features of Immune System Immune System: DefinitionDocument17 pagesGeneral Features of Immune System Immune System: Definitionمحمد نعیم اقبالNo ratings yet
- Immuno-Virology-Bacteriology Book Version 103Document332 pagesImmuno-Virology-Bacteriology Book Version 103Goose ChapmanNo ratings yet
- Pathology LO3 NotesDocument5 pagesPathology LO3 NotesT .AlainNo ratings yet
- Lecture 1 Innate ImmunityDocument8 pagesLecture 1 Innate ImmunityOfficial NgareNo ratings yet
- Blood Physiology LAb2Document8 pagesBlood Physiology LAb2DOROTA NIEOCZYMNo ratings yet
- ImmunopathologyDocument67 pagesImmunopathologyparul8attrayNo ratings yet
- Cellular and Molecular Immunology Module1: IntroductionDocument32 pagesCellular and Molecular Immunology Module1: IntroductionAygul RamankulovaNo ratings yet
- Chapter 1immunology OverviewDocument37 pagesChapter 1immunology OverviewSania HairahNo ratings yet
- Hanrayin 1Document31 pagesHanrayin 1kabulkabulovich5No ratings yet
- Hypoglycemia and Hyperglycemia GM Eng 2022Document3 pagesHypoglycemia and Hyperglycemia GM Eng 2022kabulkabulovich5No ratings yet
- Neoplasia GM Stom Eng 2023Document18 pagesNeoplasia GM Stom Eng 2023kabulkabulovich5No ratings yet
- Maternal Adaptations To Pregnancy: Gastrointestinal TractDocument21 pagesMaternal Adaptations To Pregnancy: Gastrointestinal Tractkabulkabulovich5No ratings yet
- Disseminated Intravascular Coagulation in Infants and ChildrenDocument40 pagesDisseminated Intravascular Coagulation in Infants and Childrenkabulkabulovich5No ratings yet
- Loop Diuretics: Dosing and Major Side Effects (Uptodate)Document15 pagesLoop Diuretics: Dosing and Major Side Effects (Uptodate)kabulkabulovich5No ratings yet
- Platelet Biology (Uptodate)Document31 pagesPlatelet Biology (Uptodate)kabulkabulovich5No ratings yet
- Urinary System Test BankDocument30 pagesUrinary System Test BankVinz TombocNo ratings yet
- Normal Flora and Virulent BacteriaDocument6 pagesNormal Flora and Virulent BacteriaMarwa SaadNo ratings yet
- HSC Biology Module 7Document11 pagesHSC Biology Module 7arabellatav23No ratings yet
- Brucellosis in Cattle Interim Manual For The Veterinarian & AHT - Sept2016 - SignedDocument54 pagesBrucellosis in Cattle Interim Manual For The Veterinarian & AHT - Sept2016 - SignedSuraj_Subedi100% (3)
- Introduction To Human Disease: Humberto López Castillo, MD, PHD, CPH, Cmi-SpanishDocument26 pagesIntroduction To Human Disease: Humberto López Castillo, MD, PHD, CPH, Cmi-SpanishKayla GarciaNo ratings yet
- The Allergic Response: Kathleen Nicole B. Bidua First Year ResidentDocument44 pagesThe Allergic Response: Kathleen Nicole B. Bidua First Year ResidentNicole BiduaNo ratings yet
- Immunology: Overview (Moses Franklyn Dumbuya)Document69 pagesImmunology: Overview (Moses Franklyn Dumbuya)Moses DumbuyaNo ratings yet
- Systemic Enzyme TherapyDocument10 pagesSystemic Enzyme TherapyAndrei GabrielNo ratings yet
- Artigo 8Document10 pagesArtigo 8daisyscholliersNo ratings yet
- CytokinesDocument3 pagesCytokinesTarequl Islam NishadNo ratings yet
- Pearson Science New South Wales 9 Chapter 7 Test: PlasmodiumDocument8 pagesPearson Science New South Wales 9 Chapter 7 Test: PlasmodiumAbigail ScottNo ratings yet
- Gcmaf BookDocument33 pagesGcmaf Booklennysanchez100% (1)
- Journal Reading Marsya Y Loppies 201283003Document16 pagesJournal Reading Marsya Y Loppies 201283003Marsya Yulinesia LoppiesNo ratings yet
- Test Bank For Human Physiology From Cells To Systems 7th Edition Sherwood DownloadDocument69 pagesTest Bank For Human Physiology From Cells To Systems 7th Edition Sherwood Downloadmichaelwilliamsasrfjgwoym100% (33)
- Wound MGMT Guidelines SEPTDocument93 pagesWound MGMT Guidelines SEPTYuda ArifkaNo ratings yet
- Pathophysiology of AsthmaDocument71 pagesPathophysiology of AsthmaChin ChanNo ratings yet
- Review Flash Card by Dr. Mitch Roy MaristelaDocument205 pagesReview Flash Card by Dr. Mitch Roy MaristelaMark Justin OcampoNo ratings yet
- The Inmuen Response To Surgery and TraumaDocument8 pagesThe Inmuen Response To Surgery and TraumaCorin Boice TelloNo ratings yet
- Biological Activities of Lupeol DesecurizatOKDocument21 pagesBiological Activities of Lupeol DesecurizatOKLiviu Athos TamasNo ratings yet
- The Periodontal Health Status of Invisalign Patients A Clinical StudyDocument72 pagesThe Periodontal Health Status of Invisalign Patients A Clinical Studyplayer osamaNo ratings yet
- Cytokines - IntroductionDocument2 pagesCytokines - IntroductionTra gicNo ratings yet
- Chapter 1 Basic Immunology Ppts DZ 2010Document50 pagesChapter 1 Basic Immunology Ppts DZ 2010Eng-Hussein MohamedNo ratings yet
- Inflammatory ResponseDocument7 pagesInflammatory Responseᵇᵅᵇᵞ Anden GonzagaNo ratings yet
- Case Study On LeukemiaDocument55 pagesCase Study On LeukemialicservernoidaNo ratings yet
- General Pathology CoopDocument37 pagesGeneral Pathology CoopRaymondNo ratings yet
- Liu, W., Shi, L., & Li, S. (2019) - The Immunomodulatory Effect of Alpha-Lipoic Acid in Autoimmune Diseases. BioMed Research InternationalDocument12 pagesLiu, W., Shi, L., & Li, S. (2019) - The Immunomodulatory Effect of Alpha-Lipoic Acid in Autoimmune Diseases. BioMed Research InternationalFelicia JesslynNo ratings yet
- Pathogenesis of Diabetic Atherosclerosis PDFDocument15 pagesPathogenesis of Diabetic Atherosclerosis PDFtkgoon6349No ratings yet
- MO Training ManualDocument48 pagesMO Training ManualGIST (Gujarat Institute of Science & Technology)No ratings yet
- Alrouji 2018Document47 pagesAlrouji 2018farah maulida martaNo ratings yet































































































Download as pdf or txt
You might also like
- Robbins Pathology - Chapter 3 TransDocument18 pagesRobbins Pathology - Chapter 3 Transnath nath100% (7)
- Chapter 6 - Diseases of The Immune SystemDocument3 pagesChapter 6 - Diseases of The Immune SystemArun Kumar SinghNo ratings yet
- Basic Immunology 2Document66 pagesBasic Immunology 2JabbarTapiheruNo ratings yet
- Tugas Imunologi Dasar Fast TrackDocument23 pagesTugas Imunologi Dasar Fast TrackAnditri WeningtyasNo ratings yet
- Prologue Syllabus 2008Document14 pagesPrologue Syllabus 2008Francisco Eriberto de LimaNo ratings yet
- Lecture 1 - The Immune SystemDocument29 pagesLecture 1 - The Immune SystemGana KhaledNo ratings yet
- Lecturer One Introduction To General Principles of ImmunologyDocument6 pagesLecturer One Introduction To General Principles of ImmunologyKevin KipropNo ratings yet
- 53Document29 pages53Keerthy Venthen100% (1)
- 53 PDFDocument29 pages53 PDFBrunoNo ratings yet
- 1.notes PowerpointDocument27 pages1.notes Powerpointt4ths999No ratings yet
- Basic Immunology From The Dermatologic Point of ViewDocument87 pagesBasic Immunology From The Dermatologic Point of ViewnovelraaaNo ratings yet
- Openstax - Biology2e - ch42 (VVL)Document47 pagesOpenstax - Biology2e - ch42 (VVL)ANo ratings yet
- Immune System 2018Document3 pagesImmune System 2018Luna SartoriNo ratings yet
- Chapter 6 - Diseases of The Immune SystemDocument3 pagesChapter 6 - Diseases of The Immune SystemTurinawe Bin ByensiNo ratings yet
- The Immune System: Back To The BasicsDocument37 pagesThe Immune System: Back To The BasicsHani GowaiNo ratings yet
- BCH444 PDFDocument232 pagesBCH444 PDFYaseen Ahmad100% (1)
- Topic 1,2,3 y 4Document15 pagesTopic 1,2,3 y 4Olaya De Dios HuertaNo ratings yet
- Tugas ImunilogiDocument3 pagesTugas Imunilogilapalo46No ratings yet
- Handout For Innate 1, Innate 2 and AdaptiveDocument26 pagesHandout For Innate 1, Innate 2 and Adaptive7231598No ratings yet
- Prelims - GMJ Lecture - Module 2 III DisorderDocument5 pagesPrelims - GMJ Lecture - Module 2 III DisorderjuiceNo ratings yet
- Study Guide For Introduction To Immune SystemDocument9 pagesStudy Guide For Introduction To Immune SystemOPERAñA ELLAYZA RB DECANONo ratings yet
- Immune System To Agent InfectionDocument70 pagesImmune System To Agent Infectionliza yunitaNo ratings yet
- Chapter 09. Host Response To InfectionDocument51 pagesChapter 09. Host Response To InfectionZin LimNo ratings yet
- Innate and Acquired ImmunityDocument17 pagesInnate and Acquired ImmunityPiyush KumarNo ratings yet
- IMMUNOLOGY Flash PointsDocument3 pagesIMMUNOLOGY Flash PointsHassan AhmadNo ratings yet
- The Immune System:: Innate and Acquired ImmunityDocument47 pagesThe Immune System:: Innate and Acquired ImmunityBad BunnyNo ratings yet
- ImmunologyDocument11 pagesImmunologyBiology BảoNo ratings yet
- DAMPs, PAMPs and Alarmins. All We Need To Know About DangerDocument5 pagesDAMPs, PAMPs and Alarmins. All We Need To Know About DangerjesusNo ratings yet
- Basic Immunology (Revision) : DR Z MakatiniDocument51 pagesBasic Immunology (Revision) : DR Z MakatiniEstee van WykNo ratings yet
- Immunophysiology: DR: Omer Eltahir Registrar of PathologyDocument40 pagesImmunophysiology: DR: Omer Eltahir Registrar of PathologySafa TaweelNo ratings yet
- Immunology and Serology SolutionDocument17 pagesImmunology and Serology Solutionzakaria hossainNo ratings yet
- 12 13 2006 Life Science Introduction To Immune 2006Document99 pages12 13 2006 Life Science Introduction To Immune 2006api-3696530No ratings yet
- Defence SystemDocument33 pagesDefence Systemboneheaded9No ratings yet
- Immune Response (Antibody-Antigen Reactions)Document20 pagesImmune Response (Antibody-Antigen Reactions)SimonNo ratings yet
- M2 Lecture 8 Immune ResponseDocument23 pagesM2 Lecture 8 Immune ResponseChing CHANNo ratings yet
- Ih Lec PrelimDocument24 pagesIh Lec PrelimReann SmithNo ratings yet
- NotesDocument2 pagesNotesdiceNo ratings yet
- Disease of Immune System - Siti Mirhalina 2Document25 pagesDisease of Immune System - Siti Mirhalina 2M.Satria PerdanapNo ratings yet
- Immunology Unveiled: A Comprehensive Journey through the Human Immune System: Guardians of the Body: The Unseen Heroes of ImmunityFrom EverandImmunology Unveiled: A Comprehensive Journey through the Human Immune System: Guardians of the Body: The Unseen Heroes of ImmunityNo ratings yet
- Innate ImmunityDocument16 pagesInnate ImmunityikhaNo ratings yet
- Immunity To Infections Lecture NotesDocument40 pagesImmunity To Infections Lecture Notesfredrick damianNo ratings yet
- Immunology SummaryDocument81 pagesImmunology SummaryDiaz ShehanNo ratings yet
- 2 - The Innate ImmunityDocument4 pages2 - The Innate Immunitynomore891No ratings yet
- Inflammation and Inflammatory MediatorsDocument28 pagesInflammation and Inflammatory Mediatorsla.instagramawaNo ratings yet
- Immunology: Combined Class (Mental Health Psichatry, Clinical Anastasia, & Clinical Sciences) Year 2 30/03/2022Document30 pagesImmunology: Combined Class (Mental Health Psichatry, Clinical Anastasia, & Clinical Sciences) Year 2 30/03/2022Derick NKANDU KALUBANo ratings yet
- ImunologiDocument4 pagesImunologiDhiya NabilaNo ratings yet
- Introduction To ImmunologyDocument14 pagesIntroduction To Immunologykaiyeol exoNo ratings yet
- Immunologi Klinik Ama 09102019 PDFDocument101 pagesImmunologi Klinik Ama 09102019 PDFZas VutraNo ratings yet
- Immunometabolism IDocument29 pagesImmunometabolism IMatheus TavaresNo ratings yet
- Chap 16 Part 3 Antigen-Antibody ComplexesDocument6 pagesChap 16 Part 3 Antigen-Antibody ComplexesclrssNo ratings yet
- 4 Immune DisordersDocument11 pages4 Immune DisorderscedonuliNo ratings yet
- General Features of Immune System Immune System: DefinitionDocument17 pagesGeneral Features of Immune System Immune System: Definitionمحمد نعیم اقبالNo ratings yet
- Immuno-Virology-Bacteriology Book Version 103Document332 pagesImmuno-Virology-Bacteriology Book Version 103Goose ChapmanNo ratings yet
- Pathology LO3 NotesDocument5 pagesPathology LO3 NotesT .AlainNo ratings yet
- Lecture 1 Innate ImmunityDocument8 pagesLecture 1 Innate ImmunityOfficial NgareNo ratings yet
- Blood Physiology LAb2Document8 pagesBlood Physiology LAb2DOROTA NIEOCZYMNo ratings yet
- ImmunopathologyDocument67 pagesImmunopathologyparul8attrayNo ratings yet
- Cellular and Molecular Immunology Module1: IntroductionDocument32 pagesCellular and Molecular Immunology Module1: IntroductionAygul RamankulovaNo ratings yet
- Chapter 1immunology OverviewDocument37 pagesChapter 1immunology OverviewSania HairahNo ratings yet
- Hanrayin 1Document31 pagesHanrayin 1kabulkabulovich5No ratings yet
- Hypoglycemia and Hyperglycemia GM Eng 2022Document3 pagesHypoglycemia and Hyperglycemia GM Eng 2022kabulkabulovich5No ratings yet
- Neoplasia GM Stom Eng 2023Document18 pagesNeoplasia GM Stom Eng 2023kabulkabulovich5No ratings yet
- Maternal Adaptations To Pregnancy: Gastrointestinal TractDocument21 pagesMaternal Adaptations To Pregnancy: Gastrointestinal Tractkabulkabulovich5No ratings yet
- Disseminated Intravascular Coagulation in Infants and ChildrenDocument40 pagesDisseminated Intravascular Coagulation in Infants and Childrenkabulkabulovich5No ratings yet
- Loop Diuretics: Dosing and Major Side Effects (Uptodate)Document15 pagesLoop Diuretics: Dosing and Major Side Effects (Uptodate)kabulkabulovich5No ratings yet
- Platelet Biology (Uptodate)Document31 pagesPlatelet Biology (Uptodate)kabulkabulovich5No ratings yet
- Urinary System Test BankDocument30 pagesUrinary System Test BankVinz TombocNo ratings yet
- Normal Flora and Virulent BacteriaDocument6 pagesNormal Flora and Virulent BacteriaMarwa SaadNo ratings yet
- HSC Biology Module 7Document11 pagesHSC Biology Module 7arabellatav23No ratings yet
- Brucellosis in Cattle Interim Manual For The Veterinarian & AHT - Sept2016 - SignedDocument54 pagesBrucellosis in Cattle Interim Manual For The Veterinarian & AHT - Sept2016 - SignedSuraj_Subedi100% (3)
- Introduction To Human Disease: Humberto López Castillo, MD, PHD, CPH, Cmi-SpanishDocument26 pagesIntroduction To Human Disease: Humberto López Castillo, MD, PHD, CPH, Cmi-SpanishKayla GarciaNo ratings yet
- The Allergic Response: Kathleen Nicole B. Bidua First Year ResidentDocument44 pagesThe Allergic Response: Kathleen Nicole B. Bidua First Year ResidentNicole BiduaNo ratings yet
- Immunology: Overview (Moses Franklyn Dumbuya)Document69 pagesImmunology: Overview (Moses Franklyn Dumbuya)Moses DumbuyaNo ratings yet
- Systemic Enzyme TherapyDocument10 pagesSystemic Enzyme TherapyAndrei GabrielNo ratings yet
- Artigo 8Document10 pagesArtigo 8daisyscholliersNo ratings yet
- CytokinesDocument3 pagesCytokinesTarequl Islam NishadNo ratings yet
- Pearson Science New South Wales 9 Chapter 7 Test: PlasmodiumDocument8 pagesPearson Science New South Wales 9 Chapter 7 Test: PlasmodiumAbigail ScottNo ratings yet
- Gcmaf BookDocument33 pagesGcmaf Booklennysanchez100% (1)
- Journal Reading Marsya Y Loppies 201283003Document16 pagesJournal Reading Marsya Y Loppies 201283003Marsya Yulinesia LoppiesNo ratings yet
- Test Bank For Human Physiology From Cells To Systems 7th Edition Sherwood DownloadDocument69 pagesTest Bank For Human Physiology From Cells To Systems 7th Edition Sherwood Downloadmichaelwilliamsasrfjgwoym100% (33)
- Wound MGMT Guidelines SEPTDocument93 pagesWound MGMT Guidelines SEPTYuda ArifkaNo ratings yet
- Pathophysiology of AsthmaDocument71 pagesPathophysiology of AsthmaChin ChanNo ratings yet
- Review Flash Card by Dr. Mitch Roy MaristelaDocument205 pagesReview Flash Card by Dr. Mitch Roy MaristelaMark Justin OcampoNo ratings yet
- The Inmuen Response To Surgery and TraumaDocument8 pagesThe Inmuen Response To Surgery and TraumaCorin Boice TelloNo ratings yet
- Biological Activities of Lupeol DesecurizatOKDocument21 pagesBiological Activities of Lupeol DesecurizatOKLiviu Athos TamasNo ratings yet
- The Periodontal Health Status of Invisalign Patients A Clinical StudyDocument72 pagesThe Periodontal Health Status of Invisalign Patients A Clinical Studyplayer osamaNo ratings yet
- Cytokines - IntroductionDocument2 pagesCytokines - IntroductionTra gicNo ratings yet
- Chapter 1 Basic Immunology Ppts DZ 2010Document50 pagesChapter 1 Basic Immunology Ppts DZ 2010Eng-Hussein MohamedNo ratings yet
- Inflammatory ResponseDocument7 pagesInflammatory Responseᵇᵅᵇᵞ Anden GonzagaNo ratings yet
- Case Study On LeukemiaDocument55 pagesCase Study On LeukemialicservernoidaNo ratings yet
- General Pathology CoopDocument37 pagesGeneral Pathology CoopRaymondNo ratings yet
- Liu, W., Shi, L., & Li, S. (2019) - The Immunomodulatory Effect of Alpha-Lipoic Acid in Autoimmune Diseases. BioMed Research InternationalDocument12 pagesLiu, W., Shi, L., & Li, S. (2019) - The Immunomodulatory Effect of Alpha-Lipoic Acid in Autoimmune Diseases. BioMed Research InternationalFelicia JesslynNo ratings yet
- Pathogenesis of Diabetic Atherosclerosis PDFDocument15 pagesPathogenesis of Diabetic Atherosclerosis PDFtkgoon6349No ratings yet
- MO Training ManualDocument48 pagesMO Training ManualGIST (Gujarat Institute of Science & Technology)No ratings yet
- Alrouji 2018Document47 pagesAlrouji 2018farah maulida martaNo ratings yet