Download as pdf or txt
You might also like
- Global HACCP PlanDocument65 pagesGlobal HACCP Plansyed zia ul hassan100% (3)
- SITHCCC013 Assessment 2 - Practical ObservationDocument15 pagesSITHCCC013 Assessment 2 - Practical ObservationajayNo ratings yet
- 1 Digestion in Teleost Fishes - 2Document23 pages1 Digestion in Teleost Fishes - 2Evi Nurul IhsanNo ratings yet
- SCD Food ListDocument4 pagesSCD Food ListAnuj MittalNo ratings yet
- ZaguDocument5 pagesZaguNatalie Jane Martinet100% (2)
- Sandhurst Situational AnalysisDocument35 pagesSandhurst Situational AnalysisIshanShikarkhaneNo ratings yet
- CBR Animal StructureDocument7 pagesCBR Animal StructureArifahNo ratings yet
- Disorders of The Psittacine Gastrointestinal TractDocument21 pagesDisorders of The Psittacine Gastrointestinal TractRebeca GalvãoNo ratings yet
- Rees DaviesDocument15 pagesRees DaviesAaron WoodardNo ratings yet
- The Digestive System of Fowl: VAN Practical AssignmentDocument6 pagesThe Digestive System of Fowl: VAN Practical AssignmentSampreet SaikiaNo ratings yet
- Gastropoda: Gastropoda Temporal Range: Late Cambrian-PresentDocument6 pagesGastropoda: Gastropoda Temporal Range: Late Cambrian-Presentchiluka87No ratings yet
- Da DieuDocument16 pagesDa DieuVu NguyenNo ratings yet
- The Human Vermiform AppendixDocument33 pagesThe Human Vermiform AppendixpawchanNo ratings yet
- Vertebrate DiversityDocument14 pagesVertebrate DiversityKacang PeasNo ratings yet
- Facts About LizardDocument9 pagesFacts About LizardJatin ChaudharyNo ratings yet
- Ross 2000Document40 pagesRoss 2000nicole delgadoNo ratings yet
- Cap. 02 - General Anatomy and Physiology of ReptilesDocument23 pagesCap. 02 - General Anatomy and Physiology of ReptilesNailson JúniorNo ratings yet
- Morphological Comparison of Proventricular and Gizzard in Starling Birds Sturnus Vulgaris and PigeonDocument4 pagesMorphological Comparison of Proventricular and Gizzard in Starling Birds Sturnus Vulgaris and PigeonDr. Suhaib A.H. Al-TaaiNo ratings yet
- Beltzer Et Al - Algunas Ardeidas Del Valle de Inundación Del Río ParanáDocument28 pagesBeltzer Et Al - Algunas Ardeidas Del Valle de Inundación Del Río ParanáAlejandra ArreolaNo ratings yet
- Foraging Strategies of Deep-Sea Fish: Mauchline D. GordonDocument12 pagesForaging Strategies of Deep-Sea Fish: Mauchline D. Gordonfaiz ismaNo ratings yet
- Anatomy DigestiveDocument8 pagesAnatomy DigestiveGalih Kurniawan100% (2)
- Gut MorphologyDocument27 pagesGut MorphologyrezqNo ratings yet
- Roberts T. 2017. Anatony and Physiology of The Digestive Systema of The Oarfish Regalecus Rusellii. Ichtyol Res 64. 475-477Document3 pagesRoberts T. 2017. Anatony and Physiology of The Digestive Systema of The Oarfish Regalecus Rusellii. Ichtyol Res 64. 475-477José Vélez TacuriNo ratings yet
- #2 Chapter IIDocument17 pages#2 Chapter IIAngelyn Bantilo SerronaNo ratings yet
- Oleracea Contain 13.2% Dry Matter, 15.7% Crude Protein, 5.4% Ether ExtractionDocument47 pagesOleracea Contain 13.2% Dry Matter, 15.7% Crude Protein, 5.4% Ether ExtractionJakin Aia TapanganNo ratings yet
- Digestive System of The PoultryDocument9 pagesDigestive System of The Poultrydr.mostafa.alyahyaNo ratings yet
- JejunumDocument3 pagesJejunumspiraldaoNo ratings yet
- Stomach in VertebratesDocument6 pagesStomach in VertebratesTuhinNo ratings yet
- Assignment of Diversity of AnimalDocument6 pagesAssignment of Diversity of AnimalA TNo ratings yet
- Assignment#3-Vertebral ColumnDocument5 pagesAssignment#3-Vertebral ColumnvinlavisoNo ratings yet
- Ratory Rowth, L S Ygmy ,: Labo G Reproduction and Ife Pan of The Pacific P OctopusDocument18 pagesRatory Rowth, L S Ygmy ,: Labo G Reproduction and Ife Pan of The Pacific P OctopusGadisNo ratings yet
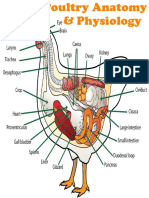
- Chicken Anatomy & PhysiologyDocument26 pagesChicken Anatomy & PhysiologySanjeev ChaudharyNo ratings yet
- Finite Element Analysis of Ungulate Jaws Can Mode PDFDocument16 pagesFinite Element Analysis of Ungulate Jaws Can Mode PDFNonoNo ratings yet
- Sustained Mass Culture Amphiascoides Atopus A Marine Harpacticoid Copepod in A Recirculating SystemDocument9 pagesSustained Mass Culture Amphiascoides Atopus A Marine Harpacticoid Copepod in A Recirculating SystemJordan IsmaelNo ratings yet
- Segers 2003 BDocument16 pagesSegers 2003 BEstefania SarangoNo ratings yet
- XXV No.3 112 215 1966 SaleDocument10 pagesXXV No.3 112 215 1966 SaleYoussefNo ratings yet
- Module 7 - Digestive SystemDocument19 pagesModule 7 - Digestive SystemkeziahlibberfNo ratings yet
- Development of The Digestive Tract of MiDocument17 pagesDevelopment of The Digestive Tract of MiLaura IbañezNo ratings yet
- 1 Lect THR 1 Intro MuscaDocument22 pages1 Lect THR 1 Intro MuscaSaadNo ratings yet
- Bird Anatomy: From Wikipedia, The Free EncyclopediaDocument12 pagesBird Anatomy: From Wikipedia, The Free Encyclopediahd_catwolfieNo ratings yet
- GastropodsDocument12 pagesGastropodsLeoregine RodriguezNo ratings yet
- Lily Issa - Bio Fun Fact! - 2Document4 pagesLily Issa - Bio Fun Fact! - 2lilyissa70No ratings yet
- MOLLUSCA 3 GastropodaDocument14 pagesMOLLUSCA 3 GastropodaCARLOSNo ratings yet
- A Novel Amniote Model of Epimorphic Regeneration The Leopard Gecko, Eublepharis MaculariusDocument25 pagesA Novel Amniote Model of Epimorphic Regeneration The Leopard Gecko, Eublepharis MaculariusSandra Clavero ClopèsNo ratings yet
- Aenigmamus (Rodentia: Diatomyidae) С Эволюционной Точки ЗренияDocument16 pagesAenigmamus (Rodentia: Diatomyidae) С Эволюционной Точки ЗренияFildzah AliyahNo ratings yet
- Comparative Vertebrate Anatomy of The Digestive System Source: Http://people - Eku.edu/ritchisong/342notes7.htmlDocument14 pagesComparative Vertebrate Anatomy of The Digestive System Source: Http://people - Eku.edu/ritchisong/342notes7.htmlYeyen M. EvoraNo ratings yet
- Anatomical Adaptations of Aquatic MammalsDocument7 pagesAnatomical Adaptations of Aquatic MammalsArturo CastroNo ratings yet
- Biology I Lecture Outline 6 Digestion & Nutrition: Pages 633 Lab 2Document17 pagesBiology I Lecture Outline 6 Digestion & Nutrition: Pages 633 Lab 2Chessking Siew HeeNo ratings yet
- Topic 8 Digestive SystemDocument10 pagesTopic 8 Digestive SystemMary Ann Morillo TenederoNo ratings yet
- Chickenanatomyphysiology 140119134735 Phpapp02Document30 pagesChickenanatomyphysiology 140119134735 Phpapp02Justine Mae Balodong BumalayNo ratings yet
- Jurnal MamaliaDocument10 pagesJurnal MamaliaClarissa B100% (1)
- 53 Publication Snakes Survive StarvationDocument10 pages53 Publication Snakes Survive StarvationÑiýāž MāłïķNo ratings yet
- Digestive System in ChordatesDocument45 pagesDigestive System in ChordatesDr. Sudesh RathodNo ratings yet
- Comparative Vertebrate Anatomy Lecture Notes - Digestive SystemDocument14 pagesComparative Vertebrate Anatomy Lecture Notes - Digestive SystemMielah Ruth100% (1)
- The Effect of Insoluble Fiber and Intermittent Feeding On Gizzard Development, Gut Motility, and Performance of Broiler ChickensDocument8 pagesThe Effect of Insoluble Fiber and Intermittent Feeding On Gizzard Development, Gut Motility, and Performance of Broiler ChickensAndre FahmiNo ratings yet
- Arthropoda Characteristics: Circulatory SystemDocument15 pagesArthropoda Characteristics: Circulatory SystemBello AjetayoNo ratings yet
- JurnalDocument4 pagesJurnalBunga Giftha NurhamsryNo ratings yet
- EntomologyDocument7 pagesEntomologymoniqueNo ratings yet
- Breeding Biology of Guppy Fish, Poecilia Reticulata: (Peters, 1859) in The LaboratoryDocument9 pagesBreeding Biology of Guppy Fish, Poecilia Reticulata: (Peters, 1859) in The LaboratoryKatamoNo ratings yet
- Breeding Biology of Guppy Fish, Poecilia Reticulata: (Peters, 1859) in The LaboratoryDocument9 pagesBreeding Biology of Guppy Fish, Poecilia Reticulata: (Peters, 1859) in The LaboratorypacifistlightNo ratings yet
- Chapter 11 - Digestive System - (Ramos, Vanny L.)Document14 pagesChapter 11 - Digestive System - (Ramos, Vanny L.)Vanny RamosNo ratings yet
- Gross and Histological Studies of Digestive Tract of Broilers During Postnatal Growth and DevelopmentDocument9 pagesGross and Histological Studies of Digestive Tract of Broilers During Postnatal Growth and DevelopmentyessiesektiNo ratings yet
- Lecture 16 Notes 2013Document4 pagesLecture 16 Notes 2013Richard HampsonNo ratings yet
- Review Article Comparison Anatomy, Physiology, A N D Biochemistry Commonly Used Laboratory AnimalsDocument30 pagesReview Article Comparison Anatomy, Physiology, A N D Biochemistry Commonly Used Laboratory AnimalsJony KechapNo ratings yet
- Horizons in Nutritional Science Carbohydrate BioavailabilityDocument11 pagesHorizons in Nutritional Science Carbohydrate BioavailabilityNurmaNo ratings yet
- Англи Хэл Тест ХариуDocument15 pagesАнгли Хэл Тест Хариуdeegijdeegij34No ratings yet
- Federal Register / Vol. 73, No. 67 / Monday, April 7, 2008 / NoticesDocument11 pagesFederal Register / Vol. 73, No. 67 / Monday, April 7, 2008 / NoticesJustia.comNo ratings yet
- Menu Journal Food Hospitality Research1Document105 pagesMenu Journal Food Hospitality Research1Juliede AlvesNo ratings yet
- Amazing Plants Teachers ManualDocument27 pagesAmazing Plants Teachers ManualSandy Hernandez100% (1)
- Co Health 10 Quarter 1Document147 pagesCo Health 10 Quarter 1Erlinda FabiaNo ratings yet
- Narrative and Pictorial Report On Gulayan Sa PaaralanDocument2 pagesNarrative and Pictorial Report On Gulayan Sa PaaralanJomarie Quirante Ü96% (50)
- Food Safety Inspection Report: Florida Department of Agriculture and Consumer Services Division of Food SafetyDocument15 pagesFood Safety Inspection Report: Florida Department of Agriculture and Consumer Services Division of Food SafetyDavid DworkNo ratings yet
- BOOK Fresh-Cut Tropical Fruits and VegetablesDocument102 pagesBOOK Fresh-Cut Tropical Fruits and VegetablesCléia Barros100% (1)
- Food Preservation MethodsDocument1 pageFood Preservation MethodsBarbara MassetteNo ratings yet
- Aboriginal Bush FoodsDocument23 pagesAboriginal Bush FoodsBeecroft Philip Adam100% (1)
- IES General Studies 2011 PDFDocument28 pagesIES General Studies 2011 PDFKshitij28No ratings yet
- Annotated BibliographyDocument4 pagesAnnotated Bibliographyapi-495053316No ratings yet
- Green Nature Triangles Project General ProposalDocument3 pagesGreen Nature Triangles Project General Proposalapi-540658681No ratings yet
- Food Adulteration: Prodyut Kumar Paul Professor, Uttar Banga Krishi ViswavidyalayaDocument25 pagesFood Adulteration: Prodyut Kumar Paul Professor, Uttar Banga Krishi ViswavidyalayaProdyut K PaulNo ratings yet
- Introduction Contains The Topics: Introduction of Seed ProductionDocument37 pagesIntroduction Contains The Topics: Introduction of Seed ProductionSheelendra Mangal BhattNo ratings yet
- Zhang 2018Document204 pagesZhang 2018Marcelo Jara FalconiNo ratings yet
- Asking and Giving OpinionsDocument6 pagesAsking and Giving OpinionsDora SilvaNo ratings yet
- Establishing Sales TargetDocument10 pagesEstablishing Sales TargetErich VirayNo ratings yet
- Synthesis: Condition and Trends in Systems and Services, Trade-Offs For Human Well-Being, and Implications For The FutureDocument12 pagesSynthesis: Condition and Trends in Systems and Services, Trade-Offs For Human Well-Being, and Implications For The FuturenmohammaNo ratings yet
- RBI Assistant Prelims 2020 The Selection PLAN 2Document75 pagesRBI Assistant Prelims 2020 The Selection PLAN 2Maddy MystroNo ratings yet
- Unhealthy BreakfastDocument2 pagesUnhealthy BreakfastOreillerNo ratings yet
- Alpaka Company Profile 2020Document25 pagesAlpaka Company Profile 2020dreamup indoNo ratings yet
- Section 5 - Students WorksheetDocument4 pagesSection 5 - Students WorksheetEsraa AhmedNo ratings yet
- FSSC AUDIT REPORT (Internal)Document3 pagesFSSC AUDIT REPORT (Internal)Md Kamruzzaman MonirNo ratings yet