Download as pdf or txt
You might also like
- Winning ResumeDocument17 pagesWinning ResumeSasi100% (4)
- Paper Envejecimiento MitocondrialDocument8 pagesPaper Envejecimiento MitocondrialCristian Angel MoyanoNo ratings yet
- Jurowski2014 Article BiologicalConsequencesOfZincDeDocument11 pagesJurowski2014 Article BiologicalConsequencesOfZincDevtimbangjrNo ratings yet
- (Andreyev Et Al) Mitochondrial Metabolism of Reactive Oxygen SpeciesDocument15 pages(Andreyev Et Al) Mitochondrial Metabolism of Reactive Oxygen SpeciesaitorNo ratings yet
- ROS y Cáncer 2020Document12 pagesROS y Cáncer 2020Maria Elena MaldonadoNo ratings yet
- ZincDocument6 pagesZincJames NorldNo ratings yet
- Zinc and Oxidative Stress - Current MechanismsDocument9 pagesZinc and Oxidative Stress - Current MechanismsNvo SpektarNo ratings yet
- Review Article Biomarkers of Oxidative Stress in Metabolic Syndrome and Associated DiseasesDocument20 pagesReview Article Biomarkers of Oxidative Stress in Metabolic Syndrome and Associated DiseasesTien TienNo ratings yet
- Omcl0204 0191Document16 pagesOmcl0204 0191Anonymous zYZi65gNo ratings yet
- Rythmic BiologyDocument2 pagesRythmic BiologyMarcelo Del PilarNo ratings yet
- Powers (2020) Exercise-Induced Oxidative Stress Friend or FoeDocument11 pagesPowers (2020) Exercise-Induced Oxidative Stress Friend or FoeBruno TeixeiraNo ratings yet
- Oxidants, Oxidative Stress and The Biology of Ageing: InsightDocument9 pagesOxidants, Oxidative Stress and The Biology of Ageing: InsightViancaGHNo ratings yet
- Mohd azri,+Journal+manager,+Chapter-3Document11 pagesMohd azri,+Journal+manager,+Chapter-3Daniela DanielaNo ratings yet
- Overview of The Antioxidant and Anti-Inflammatory Activities of Selected Plant Compounds and Their Metal Ions ComplexesDocument52 pagesOverview of The Antioxidant and Anti-Inflammatory Activities of Selected Plant Compounds and Their Metal Ions ComplexesFahmi Yusup FaisalNo ratings yet
- Oxidative Stress in Spinal Cord Injury and Antioxidant-Based InterventionDocument11 pagesOxidative Stress in Spinal Cord Injury and Antioxidant-Based InterventionZhenquan JiaNo ratings yet
- Research Notes 1994 MayDocument4 pagesResearch Notes 1994 MayRichard SmithNo ratings yet
- Cancer y EROSDocument38 pagesCancer y EROSSandra RiveraNo ratings yet
- Bowler 1994Document21 pagesBowler 1994Gheymisson BrennoNo ratings yet
- Antioxidan ReviewDocument9 pagesAntioxidan ReviewGustavo RuizNo ratings yet
- Oxidative StressDocument13 pagesOxidative StressJeniffer RafaelaNo ratings yet
- Effect of Free Radicals & Antioxidants On Oxidative Stress A ReviewDocument4 pagesEffect of Free Radicals & Antioxidants On Oxidative Stress A ReviewSeeptianMaulanaNo ratings yet
- Free Radicals, Metals and Antioxidants in Oxidative Stress-Induced CancerDocument40 pagesFree Radicals, Metals and Antioxidants in Oxidative Stress-Induced CancerAji KusumaNo ratings yet
- Critical Role of Oxidant and Antioxidant in Cancer 2168 9547.1000e110Document2 pagesCritical Role of Oxidant and Antioxidant in Cancer 2168 9547.1000e110Paulo César López BarrientosNo ratings yet
- Human Zinc DefDocument6 pagesHuman Zinc DefMiodrag MilicNo ratings yet
- Best Practice & Research Clinical Obstetrics and GynaecologyDocument13 pagesBest Practice & Research Clinical Obstetrics and Gynaecologykamis misteriNo ratings yet
- Unige 32467 Attachment01Document15 pagesUnige 32467 Attachment01Reggie SuryaNo ratings yet
- UC Irvine Previously Published WorksDocument7 pagesUC Irvine Previously Published WorksAlix AliNo ratings yet
- AterosklerosisDocument5 pagesAterosklerosisHanip NurrifqiNo ratings yet
- Taqvi 2020Document6 pagesTaqvi 2020Nguyễn Ngọc HuyềnNo ratings yet
- Fimmu 13 998059Document15 pagesFimmu 13 998059Cynthia Linniete Rodriguez MazmelaNo ratings yet
- fimmu-12-667343Document19 pagesfimmu-12-667343korosuke1920No ratings yet
- Oxidative Stress CancerDocument40 pagesOxidative Stress CancerLawrence BarreraNo ratings yet
- Antioxidants: Enzymatic and Non-Enzymatic Molecules With Antioxidant FunctionDocument4 pagesAntioxidants: Enzymatic and Non-Enzymatic Molecules With Antioxidant FunctionM Azzam SNo ratings yet
- Angeloni 2020Document28 pagesAngeloni 2020Êndel AlvesNo ratings yet
- SOD Expert Review PDFDocument11 pagesSOD Expert Review PDFMuhammad Bayu Zohari Hutagalung100% (1)
- Oxidative Stressand Redox Signalinginthe PathophysiologyofDocument8 pagesOxidative Stressand Redox Signalinginthe PathophysiologyofKIU PUBLICATION AND EXTENSIONNo ratings yet
- Oxidative stress during development Chemical-induced teratogenesisDocument18 pagesOxidative stress during development Chemical-induced teratogenesism.sonbulNo ratings yet
- Mitochondrial ROS and CancerDocument12 pagesMitochondrial ROS and CancerJaya Kunal DubeyNo ratings yet
- AVERINA Et Al, 2021 - Lactobacilos e Bifidobacterias Humano ComposiçãoDocument33 pagesAVERINA Et Al, 2021 - Lactobacilos e Bifidobacterias Humano ComposiçãoanavcmeloNo ratings yet
- Free Radicals and Human Health: July 2015Document7 pagesFree Radicals and Human Health: July 2015mabel yapuraNo ratings yet
- Oxidative Stress PDFDocument13 pagesOxidative Stress PDFFrancisco Ibañez IrribarraNo ratings yet
- Oxidative Stress, Insulin Resistance, Dyslipidemia and Type 2 Diabetes MellitusDocument26 pagesOxidative Stress, Insulin Resistance, Dyslipidemia and Type 2 Diabetes MellitusRavika Mutiara MansurNo ratings yet
- Antioxidants 12 00517 v2Document30 pagesAntioxidants 12 00517 v2luis buenaventuraNo ratings yet
- Zinc Toxicity in Humans: Jerome Nriagu, School of Public Health, University of MichiganDocument7 pagesZinc Toxicity in Humans: Jerome Nriagu, School of Public Health, University of MichiganAfshee umarNo ratings yet
- Is Young-Antioxidant in Health Disease, Dafpus 14Document11 pagesIs Young-Antioxidant in Health Disease, Dafpus 14SOPHIASTIA KUSBIANTI MHS 2017No ratings yet
- ROS and AgeingDocument18 pagesROS and AgeingAlice RizzoNo ratings yet
- Ijms 21 02173 v2 PDFDocument38 pagesIjms 21 02173 v2 PDFCarolina RicárdezNo ratings yet
- Heavy Metal Induced Oxidative Stress & Its Possible Reversal by Chelation TherapyDocument23 pagesHeavy Metal Induced Oxidative Stress & Its Possible Reversal by Chelation TherapyArnab BiswasNo ratings yet
- Redox Pioneer: Professor Joseph Loscalzo: Edox IoneerDocument8 pagesRedox Pioneer: Professor Joseph Loscalzo: Edox IoneerJosué VelázquezNo ratings yet
- 1 s2.0 S0098299711000501 MainDocument13 pages1 s2.0 S0098299711000501 Mainfilda dataNo ratings yet
- Buletin DiareDocument16 pagesBuletin DiareIka WijayaNo ratings yet
- Free Radical ParadoxesDocument2 pagesFree Radical Paradoxesngoclien93No ratings yet
- Un Objetivo para Tratar La Enfermedad de Alzheimer y El Accidente CerebrovascularDocument13 pagesUn Objetivo para Tratar La Enfermedad de Alzheimer y El Accidente CerebrovascularCristina CruzNo ratings yet
- Gosalvez 2017Document11 pagesGosalvez 2017Rakesh RamananNo ratings yet
- Antioxidantes en EmbriónDocument8 pagesAntioxidantes en EmbriónCarolina PosadaNo ratings yet
- CMCBani Bencini2012Document14 pagesCMCBani Bencini2012helidep27No ratings yet
- Reactive Oxygen Species Along With Reactive Nitrogen Species (ROS/RNS) As The Main Cause of Multiple SclerosisDocument15 pagesReactive Oxygen Species Along With Reactive Nitrogen Species (ROS/RNS) As The Main Cause of Multiple SclerosisSomayeh ZaminpiraNo ratings yet
- IN VITRO ANTIOXIDANT AND ANTIDIABETIC ACTIVITY of Plumeria PudicaDocument10 pagesIN VITRO ANTIOXIDANT AND ANTIDIABETIC ACTIVITY of Plumeria PudicaTrinity PdplNo ratings yet
- The Role of Free Radicals in Health and DiseaseDocument9 pagesThe Role of Free Radicals in Health and Diseasehumera0% (1)
- Molecular Basis of Oxidative Stress: Chemistry, Mechanisms, and Disease PathogenesisFrom EverandMolecular Basis of Oxidative Stress: Chemistry, Mechanisms, and Disease PathogenesisNo ratings yet
- Iron therapy - With Special Emphasis on Intravenous AdministrationFrom EverandIron therapy - With Special Emphasis on Intravenous AdministrationNo ratings yet
- Amish Friendship Bread RecipeDocument1 pageAmish Friendship Bread RecipeTakara Lian TaylorNo ratings yet
- Occupational TherapyDocument11 pagesOccupational TherapyYingVuong100% (1)
- Food Technology SyllabusDocument79 pagesFood Technology SyllabusJuwelRanaNo ratings yet
- Garrett PD 65001 User ManualDocument4 pagesGarrett PD 65001 User ManualFelipe MejiasNo ratings yet
- Consolidated QuestionsDocument77 pagesConsolidated QuestionsvenkatNo ratings yet
- Writing A BlogDocument5 pagesWriting A BlogJoyce SyNo ratings yet
- Tall Grass Mob StockingDocument4 pagesTall Grass Mob StockingJD_pdfNo ratings yet
- Practical Research Module 1.fDocument20 pagesPractical Research Module 1.fDianne Masapol89% (28)
- Artificial Intelligence-Opportunities and Challenges For TheDocument9 pagesArtificial Intelligence-Opportunities and Challenges For TheChris CocNo ratings yet
- Materi Tanggapan Tulisan Insan CSR Olah Karsa 2023Document18 pagesMateri Tanggapan Tulisan Insan CSR Olah Karsa 2023yantoNo ratings yet
- Hustler Magazine Inc V FalwellDocument6 pagesHustler Magazine Inc V FalwellJon Bandoma100% (1)
- TCS HR Interview QuestionsDocument10 pagesTCS HR Interview Questionsmanchiraju raj kumar100% (1)
- Criminology TermsDocument4 pagesCriminology TermsTon RJ IdoNo ratings yet
- The Guy in The GlassDocument1 pageThe Guy in The GlassAnkush AgrawalNo ratings yet
- Vantage BrunswickDocument72 pagesVantage BrunswickNathan Bukoski100% (1)
- Axis Bank - 123456763542 - Sample Statement - cc319740Document5 pagesAxis Bank - 123456763542 - Sample Statement - cc319740Seemab LatifNo ratings yet
- Navarro, Christine - Marinduque (Autosaved)Document28 pagesNavarro, Christine - Marinduque (Autosaved)CHRISTINE LOBOZANANo ratings yet
- ATF Form 4 Application For Tax Paid Transfer and Registration of FirearmDocument12 pagesATF Form 4 Application For Tax Paid Transfer and Registration of FirearmAmmoLand Shooting Sports NewsNo ratings yet
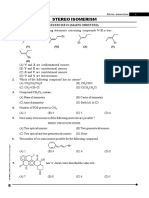
- Stereo Isomerism - (Eng)Document32 pagesStereo Isomerism - (Eng)Atharva WatekarNo ratings yet
- (Online Teaching) B1 Preliminary For Schools Writing Part 1Document6 pages(Online Teaching) B1 Preliminary For Schools Writing Part 1Florin BurugiuNo ratings yet
- File 1Document1 pageFile 1whatsappcricketstatusNo ratings yet
- Using AutoProctor - Things You - NEED - To Know - AutoProctor BlogDocument2 pagesUsing AutoProctor - Things You - NEED - To Know - AutoProctor BlogSMKN4 MATARAMNo ratings yet
- Salatin Sept03Document4 pagesSalatin Sept03api-3733859No ratings yet
- The Following Selected Transactions Were Completed PDFDocument1 pageThe Following Selected Transactions Were Completed PDFAnbu jaromia0% (1)
- Core Values Module PaoloDocument9 pagesCore Values Module PaoloLala BoraNo ratings yet
- Vaishnava Song BookDocument115 pagesVaishnava Song Bookapi-260953818No ratings yet
- Case-Digests 2 13 24 35Document8 pagesCase-Digests 2 13 24 35BingoheartNo ratings yet
- BIOGAS Problem Solving TutorialDocument16 pagesBIOGAS Problem Solving TutorialAbhishek SoniNo ratings yet
- Scania Diesel Engine Parts CatalogDocument3 pagesScania Diesel Engine Parts CatalogTarek MassimoNo ratings yet