
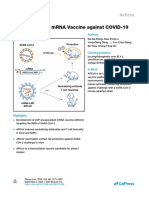
A Novel Receptor-Binding Domain (RBD) - Based Mrna Vaccine Against Sars-Cov-2
A Novel Receptor-Binding Domain (RBD) - Based Mrna Vaccine Against Sars-Cov-2
You might also like
- A Thermostable mRNA Vaccine Against COVID-19 Zhang Et - Al. 2020Document30 pagesA Thermostable mRNA Vaccine Against COVID-19 Zhang Et - Al. 2020SY LodhiNo ratings yet
- H μελέτη με θέμα: "Το RIG-I πυροδοτεί μια υπεράσπιση κατά της σηματοδότησης-άμβλυνσης αντι-SARS-CoV-2 σε ανθρώπινα πνευμονικά κύτταρα"Document60 pagesH μελέτη με θέμα: "Το RIG-I πυροδοτεί μια υπεράσπιση κατά της σηματοδότησης-άμβλυνσης αντι-SARS-CoV-2 σε ανθρώπινα πνευμονικά κύτταρα"ARISTEIDIS VIKETOSNo ratings yet
- An Antibody That Neutralizes Sars-Cov-1 and Sars-Cov-2 by Binding To A Conserved Spike Epitope Outside The Receptor Binding MotifDocument19 pagesAn Antibody That Neutralizes Sars-Cov-1 and Sars-Cov-2 by Binding To A Conserved Spike Epitope Outside The Receptor Binding Motifchato law officeNo ratings yet
- 1 s2.0 S0021925821006189 MainDocument16 pages1 s2.0 S0021925821006189 MainAngelica Maria Torregroza EspinosaNo ratings yet
- Artigo 1 - Covid-19Document3 pagesArtigo 1 - Covid-19leonardoNo ratings yet
- Versatile and Multivalent Nanobodies Efficiently Neutralize SARSCoV2ScienceDocument7 pagesVersatile and Multivalent Nanobodies Efficiently Neutralize SARSCoV2ScienceDARLENYS YADIRA MOTA MEJIANo ratings yet
- Readings On SARS-CoV-2 (Corona Virus)Document8 pagesReadings On SARS-CoV-2 (Corona Virus)negurii hearteuNo ratings yet
- JIANG 2021 (EPITOPO B) Epitope Profiling Reveals The Critical Antigenic Determinants in SARS-CoV-2 RBD-Based AntigenDocument14 pagesJIANG 2021 (EPITOPO B) Epitope Profiling Reveals The Critical Antigenic Determinants in SARS-CoV-2 RBD-Based AntigenJoão Pedro NunesNo ratings yet
- Neutralizing Antibodies Against SARS-CoV-2 CurrentDocument15 pagesNeutralizing Antibodies Against SARS-CoV-2 CurrentAntonius SimangunsongNo ratings yet
- An Alpaca Nanobody Neutralizes Sars-Cov-2 by Blocking Receptor InteractionDocument9 pagesAn Alpaca Nanobody Neutralizes Sars-Cov-2 by Blocking Receptor InteractionRicardoPeixotoNo ratings yet
- Neuropilin-1 Facilitates SARS-CoV-2 Cell Entry and Infectivity (Cantuti-Castelvetri 2020)Document7 pagesNeuropilin-1 Facilitates SARS-CoV-2 Cell Entry and Infectivity (Cantuti-Castelvetri 2020)gd_hbarNo ratings yet
- Infection Enhancing Anti Sars Cov 2 Antibodies Recognize Both The Original Wuhan D614G Strain And Delta Variants A Potential Risk For Mass Vaccination Nouara Yahi Henri Chahinian Jacques Fantini full chapter pdf docxDocument27 pagesInfection Enhancing Anti Sars Cov 2 Antibodies Recognize Both The Original Wuhan D614G Strain And Delta Variants A Potential Risk For Mass Vaccination Nouara Yahi Henri Chahinian Jacques Fantini full chapter pdf docxtolliekawann100% (7)
- Measuringtheserologic Responsetosevereacute Respiratorysyndrome Coronavirus2Document12 pagesMeasuringtheserologic Responsetosevereacute Respiratorysyndrome Coronavirus2Chris KontomarisNo ratings yet
- Lyophilization of Lipid Nanoparticle For mRNA Vaccine 1682922545Document15 pagesLyophilization of Lipid Nanoparticle For mRNA Vaccine 1682922545Piet van den OetelaarNo ratings yet
- Ab Serum CovidDocument12 pagesAb Serum CovidTania CortázarNo ratings yet
- Jurnal Sinovac 2Document6 pagesJurnal Sinovac 2Delapan SembilanNo ratings yet
- Studio Olandese Su CoronavirusDocument24 pagesStudio Olandese Su CoronavirusantiochioNo ratings yet
- Viruses: Sars-Cov-2 Spike Impairs Dna Damage Repair and Inhibits V (D) J Recombination in VitroDocument10 pagesViruses: Sars-Cov-2 Spike Impairs Dna Damage Repair and Inhibits V (D) J Recombination in Vitromuun yayo100% (1)
- Viruses: Sars-Cov-2 Spike Impairs Dna Damage Repair and Inhibits V (D) J Recombination in VitroDocument10 pagesViruses: Sars-Cov-2 Spike Impairs Dna Damage Repair and Inhibits V (D) J Recombination in VitroMohamed Fawzy mahrousNo ratings yet
- Sars-Cov-2 Can Recruit A Haem Metabolite To Evade Antibody ImmunityDocument19 pagesSars-Cov-2 Can Recruit A Haem Metabolite To Evade Antibody ImmunityShreyasri SainNo ratings yet
- WissenshaftDocument11 pagesWissenshaftkacagj bajazzoNo ratings yet
- Estudio Sobre El RemdesivirDocument28 pagesEstudio Sobre El RemdesivirA24.comNo ratings yet
- ReviewDocument8 pagesReviewIvana EstechoNo ratings yet
- 3 The Architecture of SARS CoV 2 Transcriptome 8Document19 pages3 The Architecture of SARS CoV 2 Transcriptome 8escarlet zapataNo ratings yet
- 2020 05 04 075911v1 FullDocument30 pages2020 05 04 075911v1 Fullandri priantoNo ratings yet
- Talanta: Chenchen Ge, Juan Feng, Jiaming Zhang, Kai Hu, Dou Wang, Ling Zha, Xuejuan Hu, Rongsong LiDocument9 pagesTalanta: Chenchen Ge, Juan Feng, Jiaming Zhang, Kai Hu, Dou Wang, Ling Zha, Xuejuan Hu, Rongsong LipriyaNo ratings yet
- SARS-CoV-2 IgG II ArchitectDocument10 pagesSARS-CoV-2 IgG II ArchitectcalidadlabinfectoNo ratings yet
- Protocolo - 2022Document10 pagesProtocolo - 2022Carlos Alberto Salazar DuqueNo ratings yet
- PIIS016344532300600XDocument4 pagesPIIS016344532300600Xdenemebakalim72No ratings yet
- The Pathophysiology, Diagnosis and Treatment of Corona Virus Disease 2019 (COVID-19)Document12 pagesThe Pathophysiology, Diagnosis and Treatment of Corona Virus Disease 2019 (COVID-19)GabiiGabriielaNo ratings yet
- COVID Vaccine Systematic Review Dong Dai WeiDocument14 pagesCOVID Vaccine Systematic Review Dong Dai WeiJason SnapeNo ratings yet
- Antiviral Peptides As Promising Therapeutics Against SARS-CoV 2Document8 pagesAntiviral Peptides As Promising Therapeutics Against SARS-CoV 2Stalwart sheikhNo ratings yet
- Articulo 8 CovidDocument33 pagesArticulo 8 CovidJuan OrlandoniNo ratings yet
- The SARS-CoV-2 Genome, Its Variants and Their Various Way of ImmunizationDocument7 pagesThe SARS-CoV-2 Genome, Its Variants and Their Various Way of ImmunizationInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- ZNFX1 dsRNADocument24 pagesZNFX1 dsRNAFrancesc Xavier Bofill De RosNo ratings yet
- 705 2010 Article 729Document7 pages705 2010 Article 729Lote LewaivunaNo ratings yet
- Molecular Docking Using Chimera and Autodock VinaDocument22 pagesMolecular Docking Using Chimera and Autodock VinaShisir PuriNo ratings yet
- Two Linear Epitopes On The SARS-CoV-2 SpikeDocument7 pagesTwo Linear Epitopes On The SARS-CoV-2 SpikeRheum RaponticumNo ratings yet
- Sars-Cov-2 Nsp1 Suppresses Host But Not Viral Translation Through A Bipartite MechanismDocument16 pagesSars-Cov-2 Nsp1 Suppresses Host But Not Viral Translation Through A Bipartite MechanismJanina PuenteNo ratings yet
- Journal Pre-Proof: Infection, Genetics and EvolutionDocument29 pagesJournal Pre-Proof: Infection, Genetics and EvolutionMohammed Shuaib AhmedNo ratings yet
- Crispr-Cas12 Sars Cov2Document8 pagesCrispr-Cas12 Sars Cov2Brayann InlvNo ratings yet
- Longitudinal Evaluation and Decline of Antibody Responses in Sars-Cov-2 InfectionDocument24 pagesLongitudinal Evaluation and Decline of Antibody Responses in Sars-Cov-2 InfectionCesar ValleNo ratings yet
- Novel Drugs Targeting The Sars-Cov-2/Covid-19 Machinery: Ariane Sternberg, Dwight L. Mckee and Cord NaujokatDocument11 pagesNovel Drugs Targeting The Sars-Cov-2/Covid-19 Machinery: Ariane Sternberg, Dwight L. Mckee and Cord Naujokatperi umardianaNo ratings yet
- A Prefusion SARS-CoV-2 Spike RNA Vaccine 2020.09.08.280818v1.fullDocument38 pagesA Prefusion SARS-CoV-2 Spike RNA Vaccine 2020.09.08.280818v1.fullDavid MoraNo ratings yet
- 1 s2.0 S1074761320303265 MainDocument17 pages1 s2.0 S1074761320303265 MainAleksandar DimkovskiNo ratings yet
- DiB 2019 CarvalhoDocument11 pagesDiB 2019 CarvalhoAristobolo SilvaNo ratings yet
- Processes: Trimeric Sars-Cov-2 Spike Proteins Produced From Cho Cells in Bioreactors Are High-Quality AntigensDocument11 pagesProcesses: Trimeric Sars-Cov-2 Spike Proteins Produced From Cho Cells in Bioreactors Are High-Quality AntigensHan VoNo ratings yet
- Kimia Medisinal: Oleh Kelompok 8Document16 pagesKimia Medisinal: Oleh Kelompok 8Hasryanto YantoNo ratings yet
- Zerospike Project BrochureDocument24 pagesZerospike Project Brochurestephen.cordnerNo ratings yet
- Biomedicines 10 01950Document16 pagesBiomedicines 10 01950Lija LajiNo ratings yet
- Sars-Cov-2: Phylogenetic Status, Mutations and Therapeutic Research Based On Spike ProteinDocument10 pagesSars-Cov-2: Phylogenetic Status, Mutations and Therapeutic Research Based On Spike ProteinArley GutarraNo ratings yet
- Kim Et Al., 2020Document19 pagesKim Et Al., 2020신유찬No ratings yet
- Journal Pre-Proof: Journal of Controlled ReleaseDocument17 pagesJournal Pre-Proof: Journal of Controlled ReleaseGisela GloryNo ratings yet
- Rapid Test For Detecting Neutralizing Anti RBD Antibodies PDFDocument4 pagesRapid Test For Detecting Neutralizing Anti RBD Antibodies PDFFernando GTgroupNo ratings yet
- Antigenic Characterization of The SARS-CoV-2 Omicron Subvariant BA.2.75Document26 pagesAntigenic Characterization of The SARS-CoV-2 Omicron Subvariant BA.2.75Shaviera dentiNo ratings yet
- Computers in Biology and MedicineDocument10 pagesComputers in Biology and MedicineTausif AlamNo ratings yet
- Mansfield 2004Document5 pagesMansfield 2004Nico Alexander ReyesNo ratings yet
- Sars-Cov-2 Infection Triggers Widespread Host Mrna Decay Leading To An Mrna Export BlockDocument39 pagesSars-Cov-2 Infection Triggers Widespread Host Mrna Decay Leading To An Mrna Export BlockStacey KrellerNo ratings yet
- Vaccination Versus Hybrid ImmunizationDocument9 pagesVaccination Versus Hybrid ImmunizationJovanaNo ratings yet
- Jurnal Kampen 2020Document29 pagesJurnal Kampen 2020Crosco BDLNo ratings yet
- Self-Amplifying RNA Vaccines For Infectious DiseasesDocument13 pagesSelf-Amplifying RNA Vaccines For Infectious DiseasesRicardo SousaNo ratings yet
- Adaptive Immunity To SARS-CoV-2 and COVID-19Document21 pagesAdaptive Immunity To SARS-CoV-2 and COVID-19Yuri YogyaNo ratings yet
- 089 44 Final Biologi Fasa4 DLP T4-62-84Document23 pages089 44 Final Biologi Fasa4 DLP T4-62-84BF CLNo ratings yet
- NEJMoa 2028436Document12 pagesNEJMoa 2028436arman ahdokhshNo ratings yet
- Enders 1954 - Propagation-In-Tissue-Cultures-Of-Cytopathogenic-Agents-From-Patients-With-MeaslesDocument10 pagesEnders 1954 - Propagation-In-Tissue-Cultures-Of-Cytopathogenic-Agents-From-Patients-With-MeaslesRoger TNo ratings yet
- How Effective Are COVIDDocument4 pagesHow Effective Are COVIDMARHUMA HAPSIN. ABBASNo ratings yet
- Article-2021-COVID-19-Neutralizing Antibodies Predict Disease Severity and SurvivalDocument25 pagesArticle-2021-COVID-19-Neutralizing Antibodies Predict Disease Severity and Survivalcarlos ArozamenaNo ratings yet
- Science Abm3425Document24 pagesScience Abm3425Lindomar Bonfim NobreNo ratings yet
- Corti Et Al., 2021Document38 pagesCorti Et Al., 2021LunaNo ratings yet
- Dog & Cat Import FormDocument4 pagesDog & Cat Import FormDavid Paul HensonNo ratings yet
- Anti HBeDocument6 pagesAnti HBeNaveen Kumar MNo ratings yet
- J Transci 2018 10 013Document7 pagesJ Transci 2018 10 013Clara Espinosa ArnandisNo ratings yet
- Chimeric Peptidomimetic Antibiotic Efficiently Neutralizes Lipopolysaccharides (LPS) and Bacteria-Induced Activation of RAW MacrophagesDocument9 pagesChimeric Peptidomimetic Antibiotic Efficiently Neutralizes Lipopolysaccharides (LPS) and Bacteria-Induced Activation of RAW MacrophagesKATIA PAMELA VILLAVICENCIO SANCHEZNo ratings yet
- ViruSure Brochure 2020Document20 pagesViruSure Brochure 2020joeNo ratings yet
- Aid 2011 1502Document148 pagesAid 2011 1502pasargadae_mNo ratings yet
- Jordan2017 Infectious BronchitisDocument28 pagesJordan2017 Infectious BronchitisVicente Vega SanchezNo ratings yet
- 5 Concerns About SARS-CoV2 Biology: A Call To Pause, Deliberate, and Revise Policy - by J Jay Couey - Gigaohm Biological - MediumDocument24 pages5 Concerns About SARS-CoV2 Biology: A Call To Pause, Deliberate, and Revise Policy - by J Jay Couey - Gigaohm Biological - MediumAnonymous lEJuFJMNo ratings yet
- The Journey of Antibody and Antigen Test During COVID-19 Pandemic in Indonesia, The Advantages & DisadvantagesDocument34 pagesThe Journey of Antibody and Antigen Test During COVID-19 Pandemic in Indonesia, The Advantages & DisadvantagesEldo TaufilaNo ratings yet
- Laboratory Testing For COVID-19 - Dr. Trilis Yulianti, M.kesDocument20 pagesLaboratory Testing For COVID-19 - Dr. Trilis Yulianti, M.kesYayax RakhmanNo ratings yet
- DR Robert Malone Speaks OutDocument40 pagesDR Robert Malone Speaks OutB100% (2)
- Rational Design of A Triple-Type Human Papillomavirus Vaccine by Compromising Viral-Type Speci FicityDocument15 pagesRational Design of A Triple-Type Human Papillomavirus Vaccine by Compromising Viral-Type Speci FicitySamer ShamshadNo ratings yet
- Laboratory Techniques in Rabies: Fourth EditionDocument494 pagesLaboratory Techniques in Rabies: Fourth EditionKimjhee Yang WongNo ratings yet
- Tests Krok Gen Bact VirDocument18 pagesTests Krok Gen Bact Viryasserzeddou3000No ratings yet
- Materi Pembicara Dr. TrilisDocument13 pagesMateri Pembicara Dr. TrilisDamian HamzahNo ratings yet
- Nanomaterials 11 01788 v2 5Document25 pagesNanomaterials 11 01788 v2 5Mark LaceyNo ratings yet
- jn.1 Ire CleanDocument6 pagesjn.1 Ire CleanKabakçı HüsnüNo ratings yet
- Produksi Kolostrum Antivirus Avian InfluenzaDocument11 pagesProduksi Kolostrum Antivirus Avian InfluenzaRohendi RohendiNo ratings yet
- Immune Response To HIVDocument9 pagesImmune Response To HIVTugas HeinzNo ratings yet
- Respiratory Virus InfectionsDocument9 pagesRespiratory Virus InfectionsChawki MokademNo ratings yet
























































































Download as pdf or txt
You might also like
- A Thermostable mRNA Vaccine Against COVID-19 Zhang Et - Al. 2020Document30 pagesA Thermostable mRNA Vaccine Against COVID-19 Zhang Et - Al. 2020SY LodhiNo ratings yet
- H μελέτη με θέμα: "Το RIG-I πυροδοτεί μια υπεράσπιση κατά της σηματοδότησης-άμβλυνσης αντι-SARS-CoV-2 σε ανθρώπινα πνευμονικά κύτταρα"Document60 pagesH μελέτη με θέμα: "Το RIG-I πυροδοτεί μια υπεράσπιση κατά της σηματοδότησης-άμβλυνσης αντι-SARS-CoV-2 σε ανθρώπινα πνευμονικά κύτταρα"ARISTEIDIS VIKETOSNo ratings yet
- An Antibody That Neutralizes Sars-Cov-1 and Sars-Cov-2 by Binding To A Conserved Spike Epitope Outside The Receptor Binding MotifDocument19 pagesAn Antibody That Neutralizes Sars-Cov-1 and Sars-Cov-2 by Binding To A Conserved Spike Epitope Outside The Receptor Binding Motifchato law officeNo ratings yet
- 1 s2.0 S0021925821006189 MainDocument16 pages1 s2.0 S0021925821006189 MainAngelica Maria Torregroza EspinosaNo ratings yet
- Artigo 1 - Covid-19Document3 pagesArtigo 1 - Covid-19leonardoNo ratings yet
- Versatile and Multivalent Nanobodies Efficiently Neutralize SARSCoV2ScienceDocument7 pagesVersatile and Multivalent Nanobodies Efficiently Neutralize SARSCoV2ScienceDARLENYS YADIRA MOTA MEJIANo ratings yet
- Readings On SARS-CoV-2 (Corona Virus)Document8 pagesReadings On SARS-CoV-2 (Corona Virus)negurii hearteuNo ratings yet
- JIANG 2021 (EPITOPO B) Epitope Profiling Reveals The Critical Antigenic Determinants in SARS-CoV-2 RBD-Based AntigenDocument14 pagesJIANG 2021 (EPITOPO B) Epitope Profiling Reveals The Critical Antigenic Determinants in SARS-CoV-2 RBD-Based AntigenJoão Pedro NunesNo ratings yet
- Neutralizing Antibodies Against SARS-CoV-2 CurrentDocument15 pagesNeutralizing Antibodies Against SARS-CoV-2 CurrentAntonius SimangunsongNo ratings yet
- An Alpaca Nanobody Neutralizes Sars-Cov-2 by Blocking Receptor InteractionDocument9 pagesAn Alpaca Nanobody Neutralizes Sars-Cov-2 by Blocking Receptor InteractionRicardoPeixotoNo ratings yet
- Neuropilin-1 Facilitates SARS-CoV-2 Cell Entry and Infectivity (Cantuti-Castelvetri 2020)Document7 pagesNeuropilin-1 Facilitates SARS-CoV-2 Cell Entry and Infectivity (Cantuti-Castelvetri 2020)gd_hbarNo ratings yet
- Infection Enhancing Anti Sars Cov 2 Antibodies Recognize Both The Original Wuhan D614G Strain And Delta Variants A Potential Risk For Mass Vaccination Nouara Yahi Henri Chahinian Jacques Fantini full chapter pdf docxDocument27 pagesInfection Enhancing Anti Sars Cov 2 Antibodies Recognize Both The Original Wuhan D614G Strain And Delta Variants A Potential Risk For Mass Vaccination Nouara Yahi Henri Chahinian Jacques Fantini full chapter pdf docxtolliekawann100% (7)
- Measuringtheserologic Responsetosevereacute Respiratorysyndrome Coronavirus2Document12 pagesMeasuringtheserologic Responsetosevereacute Respiratorysyndrome Coronavirus2Chris KontomarisNo ratings yet
- Lyophilization of Lipid Nanoparticle For mRNA Vaccine 1682922545Document15 pagesLyophilization of Lipid Nanoparticle For mRNA Vaccine 1682922545Piet van den OetelaarNo ratings yet
- Ab Serum CovidDocument12 pagesAb Serum CovidTania CortázarNo ratings yet
- Jurnal Sinovac 2Document6 pagesJurnal Sinovac 2Delapan SembilanNo ratings yet
- Studio Olandese Su CoronavirusDocument24 pagesStudio Olandese Su CoronavirusantiochioNo ratings yet
- Viruses: Sars-Cov-2 Spike Impairs Dna Damage Repair and Inhibits V (D) J Recombination in VitroDocument10 pagesViruses: Sars-Cov-2 Spike Impairs Dna Damage Repair and Inhibits V (D) J Recombination in Vitromuun yayo100% (1)
- Viruses: Sars-Cov-2 Spike Impairs Dna Damage Repair and Inhibits V (D) J Recombination in VitroDocument10 pagesViruses: Sars-Cov-2 Spike Impairs Dna Damage Repair and Inhibits V (D) J Recombination in VitroMohamed Fawzy mahrousNo ratings yet
- Sars-Cov-2 Can Recruit A Haem Metabolite To Evade Antibody ImmunityDocument19 pagesSars-Cov-2 Can Recruit A Haem Metabolite To Evade Antibody ImmunityShreyasri SainNo ratings yet
- WissenshaftDocument11 pagesWissenshaftkacagj bajazzoNo ratings yet
- Estudio Sobre El RemdesivirDocument28 pagesEstudio Sobre El RemdesivirA24.comNo ratings yet
- ReviewDocument8 pagesReviewIvana EstechoNo ratings yet
- 3 The Architecture of SARS CoV 2 Transcriptome 8Document19 pages3 The Architecture of SARS CoV 2 Transcriptome 8escarlet zapataNo ratings yet
- 2020 05 04 075911v1 FullDocument30 pages2020 05 04 075911v1 Fullandri priantoNo ratings yet
- Talanta: Chenchen Ge, Juan Feng, Jiaming Zhang, Kai Hu, Dou Wang, Ling Zha, Xuejuan Hu, Rongsong LiDocument9 pagesTalanta: Chenchen Ge, Juan Feng, Jiaming Zhang, Kai Hu, Dou Wang, Ling Zha, Xuejuan Hu, Rongsong LipriyaNo ratings yet
- SARS-CoV-2 IgG II ArchitectDocument10 pagesSARS-CoV-2 IgG II ArchitectcalidadlabinfectoNo ratings yet
- Protocolo - 2022Document10 pagesProtocolo - 2022Carlos Alberto Salazar DuqueNo ratings yet
- PIIS016344532300600XDocument4 pagesPIIS016344532300600Xdenemebakalim72No ratings yet
- The Pathophysiology, Diagnosis and Treatment of Corona Virus Disease 2019 (COVID-19)Document12 pagesThe Pathophysiology, Diagnosis and Treatment of Corona Virus Disease 2019 (COVID-19)GabiiGabriielaNo ratings yet
- COVID Vaccine Systematic Review Dong Dai WeiDocument14 pagesCOVID Vaccine Systematic Review Dong Dai WeiJason SnapeNo ratings yet
- Antiviral Peptides As Promising Therapeutics Against SARS-CoV 2Document8 pagesAntiviral Peptides As Promising Therapeutics Against SARS-CoV 2Stalwart sheikhNo ratings yet
- Articulo 8 CovidDocument33 pagesArticulo 8 CovidJuan OrlandoniNo ratings yet
- The SARS-CoV-2 Genome, Its Variants and Their Various Way of ImmunizationDocument7 pagesThe SARS-CoV-2 Genome, Its Variants and Their Various Way of ImmunizationInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- ZNFX1 dsRNADocument24 pagesZNFX1 dsRNAFrancesc Xavier Bofill De RosNo ratings yet
- 705 2010 Article 729Document7 pages705 2010 Article 729Lote LewaivunaNo ratings yet
- Molecular Docking Using Chimera and Autodock VinaDocument22 pagesMolecular Docking Using Chimera and Autodock VinaShisir PuriNo ratings yet
- Two Linear Epitopes On The SARS-CoV-2 SpikeDocument7 pagesTwo Linear Epitopes On The SARS-CoV-2 SpikeRheum RaponticumNo ratings yet
- Sars-Cov-2 Nsp1 Suppresses Host But Not Viral Translation Through A Bipartite MechanismDocument16 pagesSars-Cov-2 Nsp1 Suppresses Host But Not Viral Translation Through A Bipartite MechanismJanina PuenteNo ratings yet
- Journal Pre-Proof: Infection, Genetics and EvolutionDocument29 pagesJournal Pre-Proof: Infection, Genetics and EvolutionMohammed Shuaib AhmedNo ratings yet
- Crispr-Cas12 Sars Cov2Document8 pagesCrispr-Cas12 Sars Cov2Brayann InlvNo ratings yet
- Longitudinal Evaluation and Decline of Antibody Responses in Sars-Cov-2 InfectionDocument24 pagesLongitudinal Evaluation and Decline of Antibody Responses in Sars-Cov-2 InfectionCesar ValleNo ratings yet
- Novel Drugs Targeting The Sars-Cov-2/Covid-19 Machinery: Ariane Sternberg, Dwight L. Mckee and Cord NaujokatDocument11 pagesNovel Drugs Targeting The Sars-Cov-2/Covid-19 Machinery: Ariane Sternberg, Dwight L. Mckee and Cord Naujokatperi umardianaNo ratings yet
- A Prefusion SARS-CoV-2 Spike RNA Vaccine 2020.09.08.280818v1.fullDocument38 pagesA Prefusion SARS-CoV-2 Spike RNA Vaccine 2020.09.08.280818v1.fullDavid MoraNo ratings yet
- 1 s2.0 S1074761320303265 MainDocument17 pages1 s2.0 S1074761320303265 MainAleksandar DimkovskiNo ratings yet
- DiB 2019 CarvalhoDocument11 pagesDiB 2019 CarvalhoAristobolo SilvaNo ratings yet
- Processes: Trimeric Sars-Cov-2 Spike Proteins Produced From Cho Cells in Bioreactors Are High-Quality AntigensDocument11 pagesProcesses: Trimeric Sars-Cov-2 Spike Proteins Produced From Cho Cells in Bioreactors Are High-Quality AntigensHan VoNo ratings yet
- Kimia Medisinal: Oleh Kelompok 8Document16 pagesKimia Medisinal: Oleh Kelompok 8Hasryanto YantoNo ratings yet
- Zerospike Project BrochureDocument24 pagesZerospike Project Brochurestephen.cordnerNo ratings yet
- Biomedicines 10 01950Document16 pagesBiomedicines 10 01950Lija LajiNo ratings yet
- Sars-Cov-2: Phylogenetic Status, Mutations and Therapeutic Research Based On Spike ProteinDocument10 pagesSars-Cov-2: Phylogenetic Status, Mutations and Therapeutic Research Based On Spike ProteinArley GutarraNo ratings yet
- Kim Et Al., 2020Document19 pagesKim Et Al., 2020신유찬No ratings yet
- Journal Pre-Proof: Journal of Controlled ReleaseDocument17 pagesJournal Pre-Proof: Journal of Controlled ReleaseGisela GloryNo ratings yet
- Rapid Test For Detecting Neutralizing Anti RBD Antibodies PDFDocument4 pagesRapid Test For Detecting Neutralizing Anti RBD Antibodies PDFFernando GTgroupNo ratings yet
- Antigenic Characterization of The SARS-CoV-2 Omicron Subvariant BA.2.75Document26 pagesAntigenic Characterization of The SARS-CoV-2 Omicron Subvariant BA.2.75Shaviera dentiNo ratings yet
- Computers in Biology and MedicineDocument10 pagesComputers in Biology and MedicineTausif AlamNo ratings yet
- Mansfield 2004Document5 pagesMansfield 2004Nico Alexander ReyesNo ratings yet
- Sars-Cov-2 Infection Triggers Widespread Host Mrna Decay Leading To An Mrna Export BlockDocument39 pagesSars-Cov-2 Infection Triggers Widespread Host Mrna Decay Leading To An Mrna Export BlockStacey KrellerNo ratings yet
- Vaccination Versus Hybrid ImmunizationDocument9 pagesVaccination Versus Hybrid ImmunizationJovanaNo ratings yet
- Jurnal Kampen 2020Document29 pagesJurnal Kampen 2020Crosco BDLNo ratings yet
- Self-Amplifying RNA Vaccines For Infectious DiseasesDocument13 pagesSelf-Amplifying RNA Vaccines For Infectious DiseasesRicardo SousaNo ratings yet
- Adaptive Immunity To SARS-CoV-2 and COVID-19Document21 pagesAdaptive Immunity To SARS-CoV-2 and COVID-19Yuri YogyaNo ratings yet
- 089 44 Final Biologi Fasa4 DLP T4-62-84Document23 pages089 44 Final Biologi Fasa4 DLP T4-62-84BF CLNo ratings yet
- NEJMoa 2028436Document12 pagesNEJMoa 2028436arman ahdokhshNo ratings yet
- Enders 1954 - Propagation-In-Tissue-Cultures-Of-Cytopathogenic-Agents-From-Patients-With-MeaslesDocument10 pagesEnders 1954 - Propagation-In-Tissue-Cultures-Of-Cytopathogenic-Agents-From-Patients-With-MeaslesRoger TNo ratings yet
- How Effective Are COVIDDocument4 pagesHow Effective Are COVIDMARHUMA HAPSIN. ABBASNo ratings yet
- Article-2021-COVID-19-Neutralizing Antibodies Predict Disease Severity and SurvivalDocument25 pagesArticle-2021-COVID-19-Neutralizing Antibodies Predict Disease Severity and Survivalcarlos ArozamenaNo ratings yet
- Science Abm3425Document24 pagesScience Abm3425Lindomar Bonfim NobreNo ratings yet
- Corti Et Al., 2021Document38 pagesCorti Et Al., 2021LunaNo ratings yet
- Dog & Cat Import FormDocument4 pagesDog & Cat Import FormDavid Paul HensonNo ratings yet
- Anti HBeDocument6 pagesAnti HBeNaveen Kumar MNo ratings yet
- J Transci 2018 10 013Document7 pagesJ Transci 2018 10 013Clara Espinosa ArnandisNo ratings yet
- Chimeric Peptidomimetic Antibiotic Efficiently Neutralizes Lipopolysaccharides (LPS) and Bacteria-Induced Activation of RAW MacrophagesDocument9 pagesChimeric Peptidomimetic Antibiotic Efficiently Neutralizes Lipopolysaccharides (LPS) and Bacteria-Induced Activation of RAW MacrophagesKATIA PAMELA VILLAVICENCIO SANCHEZNo ratings yet
- ViruSure Brochure 2020Document20 pagesViruSure Brochure 2020joeNo ratings yet
- Aid 2011 1502Document148 pagesAid 2011 1502pasargadae_mNo ratings yet
- Jordan2017 Infectious BronchitisDocument28 pagesJordan2017 Infectious BronchitisVicente Vega SanchezNo ratings yet
- 5 Concerns About SARS-CoV2 Biology: A Call To Pause, Deliberate, and Revise Policy - by J Jay Couey - Gigaohm Biological - MediumDocument24 pages5 Concerns About SARS-CoV2 Biology: A Call To Pause, Deliberate, and Revise Policy - by J Jay Couey - Gigaohm Biological - MediumAnonymous lEJuFJMNo ratings yet
- The Journey of Antibody and Antigen Test During COVID-19 Pandemic in Indonesia, The Advantages & DisadvantagesDocument34 pagesThe Journey of Antibody and Antigen Test During COVID-19 Pandemic in Indonesia, The Advantages & DisadvantagesEldo TaufilaNo ratings yet
- Laboratory Testing For COVID-19 - Dr. Trilis Yulianti, M.kesDocument20 pagesLaboratory Testing For COVID-19 - Dr. Trilis Yulianti, M.kesYayax RakhmanNo ratings yet
- DR Robert Malone Speaks OutDocument40 pagesDR Robert Malone Speaks OutB100% (2)
- Rational Design of A Triple-Type Human Papillomavirus Vaccine by Compromising Viral-Type Speci FicityDocument15 pagesRational Design of A Triple-Type Human Papillomavirus Vaccine by Compromising Viral-Type Speci FicitySamer ShamshadNo ratings yet
- Laboratory Techniques in Rabies: Fourth EditionDocument494 pagesLaboratory Techniques in Rabies: Fourth EditionKimjhee Yang WongNo ratings yet
- Tests Krok Gen Bact VirDocument18 pagesTests Krok Gen Bact Viryasserzeddou3000No ratings yet
- Materi Pembicara Dr. TrilisDocument13 pagesMateri Pembicara Dr. TrilisDamian HamzahNo ratings yet
- Nanomaterials 11 01788 v2 5Document25 pagesNanomaterials 11 01788 v2 5Mark LaceyNo ratings yet
- jn.1 Ire CleanDocument6 pagesjn.1 Ire CleanKabakçı HüsnüNo ratings yet
- Produksi Kolostrum Antivirus Avian InfluenzaDocument11 pagesProduksi Kolostrum Antivirus Avian InfluenzaRohendi RohendiNo ratings yet
- Immune Response To HIVDocument9 pagesImmune Response To HIVTugas HeinzNo ratings yet
- Respiratory Virus InfectionsDocument9 pagesRespiratory Virus InfectionsChawki MokademNo ratings yet