
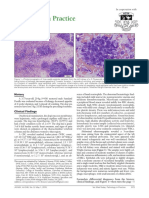
Weiser Kociba 1983 Erythrocyte Macrocytosis in Feline Leukemia Virus Associated Anemia
Weiser Kociba 1983 Erythrocyte Macrocytosis in Feline Leukemia Virus Associated Anemia
You might also like
- COURSE PLAN Child Health NursingDocument11 pagesCOURSE PLAN Child Health NursingSanthosh.S.U33% (3)
- Blood Group in AnimalDocument15 pagesBlood Group in AnimalAashish Dhakal100% (3)
- Schizophrenia PPT For EditDocument75 pagesSchizophrenia PPT For EditCharline A. Radislao100% (1)
- S. Connolly Mini Guide Astral Temple ModifiedDocument31 pagesS. Connolly Mini Guide Astral Temple Modifiedbuddaofdestruction100% (10)
- Piggery DocxdDocument7 pagesPiggery DocxdRamesiz Piad100% (2)
- Veterinary Clinical PathologyDocument15 pagesVeterinary Clinical Pathologypand00raNo ratings yet
- Plaquetas ErlichiaDocument6 pagesPlaquetas ErlichiaDIANA PATRICIA MOLANO PEREZNo ratings yet
- j.1939-1676.1991.tb03132.x InglesDocument5 pagesj.1939-1676.1991.tb03132.x InglesEdù Ccarhuani QuispeNo ratings yet
- Hoff Et Al 1985 An Appraisal of Bone Marrow Biopsy in Assessment of Sick DogsDocument9 pagesHoff Et Al 1985 An Appraisal of Bone Marrow Biopsy in Assessment of Sick DogsamnicolNo ratings yet
- Paper ADocument7 pagesPaper AdeltanueveNo ratings yet
- Approach To Feline Anaemia Cases Severine TaskerDocument7 pagesApproach To Feline Anaemia Cases Severine TaskerTomiOngkoWijayaSigalinggingNo ratings yet
- Lfurbot,+aabp 2000 v34 2 005 HematologicalAbnormalitiesDocument6 pagesLfurbot,+aabp 2000 v34 2 005 HematologicalAbnormalitiesLaura RosasNo ratings yet
- 10 Transfusion GatosDocument4 pages10 Transfusion GatosJoce MaripanguiNo ratings yet
- Pathogenesis Canine Parvovirus Enteritis: The Importance of ViremiaDocument12 pagesPathogenesis Canine Parvovirus Enteritis: The Importance of ViremiaDiana OntiverosNo ratings yet
- Theileriosis and Babesiosis in Cattle Haemogram and Some Biochemical ParametersDocument9 pagesTheileriosis and Babesiosis in Cattle Haemogram and Some Biochemical ParametersHusseinNo ratings yet
- S2 3Document10 pagesS2 3Viviana MontenegroNo ratings yet
- Veterinary Clinical Pathology Vol 40Document2 pagesVeterinary Clinical Pathology Vol 40Zbysek Vichta jr.No ratings yet
- VCP3104 167-182Document16 pagesVCP3104 167-182marceldelf355No ratings yet
- Abstract AllDocument48 pagesAbstract Allyomna kilanyNo ratings yet
- Vet Pathol 2011 2011 ACVP Annual Meeting E1 E51Document52 pagesVet Pathol 2011 2011 ACVP Annual Meeting E1 E51Haroon RashidNo ratings yet
- CANINE-Hematologic and Serum Biochemical Reference Values in Retired GreyhoundsDocument6 pagesCANINE-Hematologic and Serum Biochemical Reference Values in Retired Greyhoundstaner_soysurenNo ratings yet
- Javma-Javma.2001.218.1590 2Document3 pagesJavma-Javma.2001.218.1590 2Erhan ErtuçNo ratings yet
- Veterinary Clinical Pathol - 2022 - Frye - Complete Blood Count and Biochemistry Reference Intervals For Healthy Adult-2Document7 pagesVeterinary Clinical Pathol - 2022 - Frye - Complete Blood Count and Biochemistry Reference Intervals For Healthy Adult-2LeandroNo ratings yet
- Concentraciones Sericas de Eritropoyetina en Perro y Gatos Anemicos y PolicitemicosDocument8 pagesConcentraciones Sericas de Eritropoyetina en Perro y Gatos Anemicos y PolicitemicosZoleida BastidasNo ratings yet
- Kvitko-White Et Al-2013-Journal of Veterinary Internal MedicineDocument7 pagesKvitko-White Et Al-2013-Journal of Veterinary Internal MedicineCabinet VeterinarNo ratings yet
- Jurnal Malaria TesDocument5 pagesJurnal Malaria TesMaya RustamNo ratings yet
- Case StudiesDocument56 pagesCase StudiesMayra F. MontielNo ratings yet
- Salmonella Enterica Serotype I 4, 5, 12:-: 1,2. This Is The First Report of Acute HepaticDocument12 pagesSalmonella Enterica Serotype I 4, 5, 12:-: 1,2. This Is The First Report of Acute HepaticOktavia firnandaNo ratings yet
- A Case Report of Mycoplasma Wenyonii Associated Immune-Mediated Haemolytic Anaemia in A Dairy CowDocument8 pagesA Case Report of Mycoplasma Wenyonii Associated Immune-Mediated Haemolytic Anaemia in A Dairy CowputuNo ratings yet
- Haemobartonellosis in A Domestic Cat in Indonesia: A Case StudyDocument4 pagesHaemobartonellosis in A Domestic Cat in Indonesia: A Case StudyBima Ary WibowoNo ratings yet
- (Bravo-Ramos) Manuscript - Tracked - VersionDocument12 pages(Bravo-Ramos) Manuscript - Tracked - Versionjose bravoNo ratings yet
- Clinicopathological Studies On Camel Theileriosis: Shereen Youssef El-Sayed Abd-Elmoteleb YoussefDocument20 pagesClinicopathological Studies On Camel Theileriosis: Shereen Youssef El-Sayed Abd-Elmoteleb YoussefHasn FXNo ratings yet
- Seminario 19 - Leptos - IDocument5 pagesSeminario 19 - Leptos - IfelipeNo ratings yet
- A Retrospective Evaluation of Phenobarbital-Induced Hematologic Changes in 69 CatsDocument6 pagesA Retrospective Evaluation of Phenobarbital-Induced Hematologic Changes in 69 CatsAkın SeverNo ratings yet
- Journal of Bacteriology 1967 Gordee 6.fullDocument7 pagesJournal of Bacteriology 1967 Gordee 6.fullProma GurNo ratings yet
- Sismex ManualDocument11 pagesSismex ManualDigo SomengNo ratings yet
- FR2 PDFDocument4 pagesFR2 PDFRaphael Joshua De GuzmanNo ratings yet
- Canine Coronavirus-Associated Puppy Mortality Without Evidence of Concurrent Canine Parvovirus InfectionDocument5 pagesCanine Coronavirus-Associated Puppy Mortality Without Evidence of Concurrent Canine Parvovirus Infectionhein heinNo ratings yet
- Admin,+4 Red Cell Alloimmunization in MT ThalassemiaMajor Pts1Document4 pagesAdmin,+4 Red Cell Alloimmunization in MT ThalassemiaMajor Pts1Shujat RazaqNo ratings yet
- Biochemical, Hematological, and Electrocardiographic Changes in Buffaloes Naturally Infected WithDocument6 pagesBiochemical, Hematological, and Electrocardiographic Changes in Buffaloes Naturally Infected WithKamilahNo ratings yet
- 104063870001200209Document3 pages104063870001200209Kent Solis VicenteNo ratings yet
- Small Animal Transfusion Medicine - 2020 - VSPDocument12 pagesSmall Animal Transfusion Medicine - 2020 - VSPTactvisNo ratings yet
- Increased Erythrocytic Osmotic Fragility in Anemic Domestic Shorthair and Purebred CatsDocument20 pagesIncreased Erythrocytic Osmotic Fragility in Anemic Domestic Shorthair and Purebred CatsThành Nguyễn VănNo ratings yet
- PNAS 2000 Elder 2999 3003Document5 pagesPNAS 2000 Elder 2999 3003haddig8No ratings yet
- Rabbit Clinical Pathology - PMCDocument20 pagesRabbit Clinical Pathology - PMCmflorentinNo ratings yet
- An Introduction To Blood Groups and Blood Transfusion in Domestic AnimalsDocument6 pagesAn Introduction To Blood Groups and Blood Transfusion in Domestic Animalsshada jrjaNo ratings yet
- VeterinerDocument47 pagesVeterinerSinthia Medchera ImmauliateNo ratings yet
- Altered Blood Procalcitonin, C-Reactive Protein, and Leucocytes Count in Association With Canine Parvovirus (CPV) EnteritisDocument5 pagesAltered Blood Procalcitonin, C-Reactive Protein, and Leucocytes Count in Association With Canine Parvovirus (CPV) EnteritisStéfano Celis ConsiglieriNo ratings yet
- Anemia of Chronic Renal Failure in DogsDocument7 pagesAnemia of Chronic Renal Failure in Dogsfawzi badiNo ratings yet
- Pericardial Effusion in A DogDocument4 pagesPericardial Effusion in A DogLinda Lineros SilvaNo ratings yet
- Acute B-Lymphoid Leukemia in A Mare: A Case Report: B. Bezdekova, M. Faldyna, O. Zapletal, P. JahnDocument7 pagesAcute B-Lymphoid Leukemia in A Mare: A Case Report: B. Bezdekova, M. Faldyna, O. Zapletal, P. JahnRaisis Farah DzakiyyahNo ratings yet
- Use of Peripheral Blood Leukocytes in The Study of Cell-Mediated Immunity in Bovine Anaplasmosis - A ReviewDocument5 pagesUse of Peripheral Blood Leukocytes in The Study of Cell-Mediated Immunity in Bovine Anaplasmosis - A Reviewyudhi arjentiniaNo ratings yet
- Saliva. Detection of Feline Leukemia Virus Infection In: 1987, 25 (5) :827. H Lutz and O JarrettDocument6 pagesSaliva. Detection of Feline Leukemia Virus Infection In: 1987, 25 (5) :827. H Lutz and O JarrettNatAsyaNo ratings yet
- 404-Article Text-2152-1-10-20220307Document3 pages404-Article Text-2152-1-10-20220307nadirhussainbuzdarNo ratings yet
- Neal, 1984 PDFDocument5 pagesNeal, 1984 PDFGutierrez Guarnizo SneiderNo ratings yet
- Veterinary Clinical Pathol - 2023 - Dohány - A Retrospective Evaluation of Phenobarbital Induced Hematologic Changes in 69Document6 pagesVeterinary Clinical Pathol - 2023 - Dohány - A Retrospective Evaluation of Phenobarbital Induced Hematologic Changes in 69Lorena TomoiagăNo ratings yet
- 38 Full PDFDocument7 pages38 Full PDFfarah rachmahNo ratings yet
- 47 Pathology in PracticeDocument3 pages47 Pathology in PracticeCarlos Alberto Chaves VelasquezNo ratings yet
- Canine Ehrlichiosis - Dr. Jibachha Sah, M.V.SC, LecturerDocument34 pagesCanine Ehrlichiosis - Dr. Jibachha Sah, M.V.SC, LecturerJibachha SahNo ratings yet
- Interpretation of The HemogramDocument5 pagesInterpretation of The HemogramMargarita MujicaNo ratings yet
- Patologia Da Amiloidose em Ovinos e CaprinosDocument10 pagesPatologia Da Amiloidose em Ovinos e Caprinosandersonviana.medvetNo ratings yet
- Hematology Romania-1Document88 pagesHematology Romania-1Ştefania MafteiNo ratings yet
- Key Questions in Clinical Farm Animal Medicine, Volume 3: Therapy, Prevention and Control of DiseaseFrom EverandKey Questions in Clinical Farm Animal Medicine, Volume 3: Therapy, Prevention and Control of DiseaseTanmoy RanaNo ratings yet
- Wavy Changes in The Whiskers of Domestic Cats Are Correlated With Feline Leukemia Virus InfectionDocument7 pagesWavy Changes in The Whiskers of Domestic Cats Are Correlated With Feline Leukemia Virus InfectionCamila QuidelNo ratings yet
- Veterinary Internal Medicne - 2009 - Gleich - Hematology and Serum Biochemistry of Feline Immunodeficiency Virus InfectedDocument7 pagesVeterinary Internal Medicne - 2009 - Gleich - Hematology and Serum Biochemistry of Feline Immunodeficiency Virus InfectedCamila QuidelNo ratings yet
- Javma-Javma 228 3 371Document6 pagesJavma-Javma 228 3 371Camila QuidelNo ratings yet
- Veterinary Internal Medicne - 2017 - Levy - Performance of 4 Point of Care Screening Tests For Feline Leukemia Virus andDocument6 pagesVeterinary Internal Medicne - 2017 - Levy - Performance of 4 Point of Care Screening Tests For Feline Leukemia Virus andCamila QuidelNo ratings yet
- Dunham 2008Document23 pagesDunham 2008Camila QuidelNo ratings yet
- Estillero, Mari Antoinette - SPED 104 - Output Week 1 and 2Document4 pagesEstillero, Mari Antoinette - SPED 104 - Output Week 1 and 2Mari Antoinette EstilleroNo ratings yet
- OlivierDocument481 pagesOlivierVetriSelvan Nagarajan0% (1)
- A Case of Mitral StenosisDocument29 pagesA Case of Mitral StenosisShadab KamalNo ratings yet
- Drug Profile: C H FN ODocument2 pagesDrug Profile: C H FN Ohamed razaNo ratings yet
- Nutrition Concepts and Controversies 14Th Edition Sizer Test Bank Full Chapter PDFDocument53 pagesNutrition Concepts and Controversies 14Th Edition Sizer Test Bank Full Chapter PDFinlawbast.ileir100% (12)
- A Guide To Performing Quality Assured Diagnostic SpirometryDocument28 pagesA Guide To Performing Quality Assured Diagnostic SpirometryLaura MoiseNo ratings yet
- Genetic Factors Associated With Periodontal DiseasesDocument50 pagesGenetic Factors Associated With Periodontal DiseasesDrKrishna DasNo ratings yet
- Discovery and Development of Nucleoside and Nucleotide Reverse-Transcriptase InhibitorsDocument8 pagesDiscovery and Development of Nucleoside and Nucleotide Reverse-Transcriptase Inhibitorsolivia523No ratings yet
- 2016 ATC Index With DDDDocument317 pages2016 ATC Index With DDDSi CentilNo ratings yet
- TechTalk - August2010 Clump in EDTA Tubes PDFDocument2 pagesTechTalk - August2010 Clump in EDTA Tubes PDFARIF AHAMMED PNo ratings yet
- Brachycera 08Document27 pagesBrachycera 08Agustin IskandarNo ratings yet
- Lymphatic Drainage of Head and NeckDocument19 pagesLymphatic Drainage of Head and NeckKelly Yeow100% (1)
- Occupational DiseasesDocument10 pagesOccupational DiseasesAbhishek tiwariNo ratings yet
- E-Patients White PaperDocument126 pagesE-Patients White PaperPatients Know BestNo ratings yet
- Nursing Care Plan Sheet SampleDocument3 pagesNursing Care Plan Sheet Sampledbryant0101100% (5)
- Web 1 - Dr. Wahyuni Indawati, Sp. A (K) - Diagnosis and Management of Allergic Rhinitis in ChildrenDocument27 pagesWeb 1 - Dr. Wahyuni Indawati, Sp. A (K) - Diagnosis and Management of Allergic Rhinitis in Childrenbungkang elsaNo ratings yet
- Dog Parks in Hume City - Feasibility StudyDocument32 pagesDog Parks in Hume City - Feasibility Studykd.prawestiNo ratings yet
- Chemotherapy - Alkylating AgentsDocument33 pagesChemotherapy - Alkylating AgentsNolan100% (1)
- Homosexuality: A History (From Ancient Greece To Gay Liberation) Vern L. BulloughDocument209 pagesHomosexuality: A History (From Ancient Greece To Gay Liberation) Vern L. Bulloughhugoprais100% (1)
- Dea 2Document15 pagesDea 2Dea Hadfina Kartika SariNo ratings yet
- Management of Alveolar Ridge ResorptionDocument29 pagesManagement of Alveolar Ridge Resorptionفواز نميرNo ratings yet
- Principles of Medical Technology Practice 1 Midterm TranseesDocument19 pagesPrinciples of Medical Technology Practice 1 Midterm TranseesSean Rafael LedesmaNo ratings yet
- CVS Specialty Pharmacy Distribution Drug ListDocument19 pagesCVS Specialty Pharmacy Distribution Drug Listdp4mNo ratings yet
- Gadtc PHCDocument246 pagesGadtc PHCshawnNo ratings yet
- The Respiratory System I.: Subject: Subject TeacherDocument4 pagesThe Respiratory System I.: Subject: Subject TeacherHyacinth FarinasNo ratings yet
- Down's Syndrome Invetigsatory ProjectDocument23 pagesDown's Syndrome Invetigsatory ProjectKrithishNo ratings yet





























































































Download as pdf or txt
You might also like
- COURSE PLAN Child Health NursingDocument11 pagesCOURSE PLAN Child Health NursingSanthosh.S.U33% (3)
- Blood Group in AnimalDocument15 pagesBlood Group in AnimalAashish Dhakal100% (3)
- Schizophrenia PPT For EditDocument75 pagesSchizophrenia PPT For EditCharline A. Radislao100% (1)
- S. Connolly Mini Guide Astral Temple ModifiedDocument31 pagesS. Connolly Mini Guide Astral Temple Modifiedbuddaofdestruction100% (10)
- Piggery DocxdDocument7 pagesPiggery DocxdRamesiz Piad100% (2)
- Veterinary Clinical PathologyDocument15 pagesVeterinary Clinical Pathologypand00raNo ratings yet
- Plaquetas ErlichiaDocument6 pagesPlaquetas ErlichiaDIANA PATRICIA MOLANO PEREZNo ratings yet
- j.1939-1676.1991.tb03132.x InglesDocument5 pagesj.1939-1676.1991.tb03132.x InglesEdù Ccarhuani QuispeNo ratings yet
- Hoff Et Al 1985 An Appraisal of Bone Marrow Biopsy in Assessment of Sick DogsDocument9 pagesHoff Et Al 1985 An Appraisal of Bone Marrow Biopsy in Assessment of Sick DogsamnicolNo ratings yet
- Paper ADocument7 pagesPaper AdeltanueveNo ratings yet
- Approach To Feline Anaemia Cases Severine TaskerDocument7 pagesApproach To Feline Anaemia Cases Severine TaskerTomiOngkoWijayaSigalinggingNo ratings yet
- Lfurbot,+aabp 2000 v34 2 005 HematologicalAbnormalitiesDocument6 pagesLfurbot,+aabp 2000 v34 2 005 HematologicalAbnormalitiesLaura RosasNo ratings yet
- 10 Transfusion GatosDocument4 pages10 Transfusion GatosJoce MaripanguiNo ratings yet
- Pathogenesis Canine Parvovirus Enteritis: The Importance of ViremiaDocument12 pagesPathogenesis Canine Parvovirus Enteritis: The Importance of ViremiaDiana OntiverosNo ratings yet
- Theileriosis and Babesiosis in Cattle Haemogram and Some Biochemical ParametersDocument9 pagesTheileriosis and Babesiosis in Cattle Haemogram and Some Biochemical ParametersHusseinNo ratings yet
- S2 3Document10 pagesS2 3Viviana MontenegroNo ratings yet
- Veterinary Clinical Pathology Vol 40Document2 pagesVeterinary Clinical Pathology Vol 40Zbysek Vichta jr.No ratings yet
- VCP3104 167-182Document16 pagesVCP3104 167-182marceldelf355No ratings yet
- Abstract AllDocument48 pagesAbstract Allyomna kilanyNo ratings yet
- Vet Pathol 2011 2011 ACVP Annual Meeting E1 E51Document52 pagesVet Pathol 2011 2011 ACVP Annual Meeting E1 E51Haroon RashidNo ratings yet
- CANINE-Hematologic and Serum Biochemical Reference Values in Retired GreyhoundsDocument6 pagesCANINE-Hematologic and Serum Biochemical Reference Values in Retired Greyhoundstaner_soysurenNo ratings yet
- Javma-Javma.2001.218.1590 2Document3 pagesJavma-Javma.2001.218.1590 2Erhan ErtuçNo ratings yet
- Veterinary Clinical Pathol - 2022 - Frye - Complete Blood Count and Biochemistry Reference Intervals For Healthy Adult-2Document7 pagesVeterinary Clinical Pathol - 2022 - Frye - Complete Blood Count and Biochemistry Reference Intervals For Healthy Adult-2LeandroNo ratings yet
- Concentraciones Sericas de Eritropoyetina en Perro y Gatos Anemicos y PolicitemicosDocument8 pagesConcentraciones Sericas de Eritropoyetina en Perro y Gatos Anemicos y PolicitemicosZoleida BastidasNo ratings yet
- Kvitko-White Et Al-2013-Journal of Veterinary Internal MedicineDocument7 pagesKvitko-White Et Al-2013-Journal of Veterinary Internal MedicineCabinet VeterinarNo ratings yet
- Jurnal Malaria TesDocument5 pagesJurnal Malaria TesMaya RustamNo ratings yet
- Case StudiesDocument56 pagesCase StudiesMayra F. MontielNo ratings yet
- Salmonella Enterica Serotype I 4, 5, 12:-: 1,2. This Is The First Report of Acute HepaticDocument12 pagesSalmonella Enterica Serotype I 4, 5, 12:-: 1,2. This Is The First Report of Acute HepaticOktavia firnandaNo ratings yet
- A Case Report of Mycoplasma Wenyonii Associated Immune-Mediated Haemolytic Anaemia in A Dairy CowDocument8 pagesA Case Report of Mycoplasma Wenyonii Associated Immune-Mediated Haemolytic Anaemia in A Dairy CowputuNo ratings yet
- Haemobartonellosis in A Domestic Cat in Indonesia: A Case StudyDocument4 pagesHaemobartonellosis in A Domestic Cat in Indonesia: A Case StudyBima Ary WibowoNo ratings yet
- (Bravo-Ramos) Manuscript - Tracked - VersionDocument12 pages(Bravo-Ramos) Manuscript - Tracked - Versionjose bravoNo ratings yet
- Clinicopathological Studies On Camel Theileriosis: Shereen Youssef El-Sayed Abd-Elmoteleb YoussefDocument20 pagesClinicopathological Studies On Camel Theileriosis: Shereen Youssef El-Sayed Abd-Elmoteleb YoussefHasn FXNo ratings yet
- Seminario 19 - Leptos - IDocument5 pagesSeminario 19 - Leptos - IfelipeNo ratings yet
- A Retrospective Evaluation of Phenobarbital-Induced Hematologic Changes in 69 CatsDocument6 pagesA Retrospective Evaluation of Phenobarbital-Induced Hematologic Changes in 69 CatsAkın SeverNo ratings yet
- Journal of Bacteriology 1967 Gordee 6.fullDocument7 pagesJournal of Bacteriology 1967 Gordee 6.fullProma GurNo ratings yet
- Sismex ManualDocument11 pagesSismex ManualDigo SomengNo ratings yet
- FR2 PDFDocument4 pagesFR2 PDFRaphael Joshua De GuzmanNo ratings yet
- Canine Coronavirus-Associated Puppy Mortality Without Evidence of Concurrent Canine Parvovirus InfectionDocument5 pagesCanine Coronavirus-Associated Puppy Mortality Without Evidence of Concurrent Canine Parvovirus Infectionhein heinNo ratings yet
- Admin,+4 Red Cell Alloimmunization in MT ThalassemiaMajor Pts1Document4 pagesAdmin,+4 Red Cell Alloimmunization in MT ThalassemiaMajor Pts1Shujat RazaqNo ratings yet
- Biochemical, Hematological, and Electrocardiographic Changes in Buffaloes Naturally Infected WithDocument6 pagesBiochemical, Hematological, and Electrocardiographic Changes in Buffaloes Naturally Infected WithKamilahNo ratings yet
- 104063870001200209Document3 pages104063870001200209Kent Solis VicenteNo ratings yet
- Small Animal Transfusion Medicine - 2020 - VSPDocument12 pagesSmall Animal Transfusion Medicine - 2020 - VSPTactvisNo ratings yet
- Increased Erythrocytic Osmotic Fragility in Anemic Domestic Shorthair and Purebred CatsDocument20 pagesIncreased Erythrocytic Osmotic Fragility in Anemic Domestic Shorthair and Purebred CatsThành Nguyễn VănNo ratings yet
- PNAS 2000 Elder 2999 3003Document5 pagesPNAS 2000 Elder 2999 3003haddig8No ratings yet
- Rabbit Clinical Pathology - PMCDocument20 pagesRabbit Clinical Pathology - PMCmflorentinNo ratings yet
- An Introduction To Blood Groups and Blood Transfusion in Domestic AnimalsDocument6 pagesAn Introduction To Blood Groups and Blood Transfusion in Domestic Animalsshada jrjaNo ratings yet
- VeterinerDocument47 pagesVeterinerSinthia Medchera ImmauliateNo ratings yet
- Altered Blood Procalcitonin, C-Reactive Protein, and Leucocytes Count in Association With Canine Parvovirus (CPV) EnteritisDocument5 pagesAltered Blood Procalcitonin, C-Reactive Protein, and Leucocytes Count in Association With Canine Parvovirus (CPV) EnteritisStéfano Celis ConsiglieriNo ratings yet
- Anemia of Chronic Renal Failure in DogsDocument7 pagesAnemia of Chronic Renal Failure in Dogsfawzi badiNo ratings yet
- Pericardial Effusion in A DogDocument4 pagesPericardial Effusion in A DogLinda Lineros SilvaNo ratings yet
- Acute B-Lymphoid Leukemia in A Mare: A Case Report: B. Bezdekova, M. Faldyna, O. Zapletal, P. JahnDocument7 pagesAcute B-Lymphoid Leukemia in A Mare: A Case Report: B. Bezdekova, M. Faldyna, O. Zapletal, P. JahnRaisis Farah DzakiyyahNo ratings yet
- Use of Peripheral Blood Leukocytes in The Study of Cell-Mediated Immunity in Bovine Anaplasmosis - A ReviewDocument5 pagesUse of Peripheral Blood Leukocytes in The Study of Cell-Mediated Immunity in Bovine Anaplasmosis - A Reviewyudhi arjentiniaNo ratings yet
- Saliva. Detection of Feline Leukemia Virus Infection In: 1987, 25 (5) :827. H Lutz and O JarrettDocument6 pagesSaliva. Detection of Feline Leukemia Virus Infection In: 1987, 25 (5) :827. H Lutz and O JarrettNatAsyaNo ratings yet
- 404-Article Text-2152-1-10-20220307Document3 pages404-Article Text-2152-1-10-20220307nadirhussainbuzdarNo ratings yet
- Neal, 1984 PDFDocument5 pagesNeal, 1984 PDFGutierrez Guarnizo SneiderNo ratings yet
- Veterinary Clinical Pathol - 2023 - Dohány - A Retrospective Evaluation of Phenobarbital Induced Hematologic Changes in 69Document6 pagesVeterinary Clinical Pathol - 2023 - Dohány - A Retrospective Evaluation of Phenobarbital Induced Hematologic Changes in 69Lorena TomoiagăNo ratings yet
- 38 Full PDFDocument7 pages38 Full PDFfarah rachmahNo ratings yet
- 47 Pathology in PracticeDocument3 pages47 Pathology in PracticeCarlos Alberto Chaves VelasquezNo ratings yet
- Canine Ehrlichiosis - Dr. Jibachha Sah, M.V.SC, LecturerDocument34 pagesCanine Ehrlichiosis - Dr. Jibachha Sah, M.V.SC, LecturerJibachha SahNo ratings yet
- Interpretation of The HemogramDocument5 pagesInterpretation of The HemogramMargarita MujicaNo ratings yet
- Patologia Da Amiloidose em Ovinos e CaprinosDocument10 pagesPatologia Da Amiloidose em Ovinos e Caprinosandersonviana.medvetNo ratings yet
- Hematology Romania-1Document88 pagesHematology Romania-1Ştefania MafteiNo ratings yet
- Key Questions in Clinical Farm Animal Medicine, Volume 3: Therapy, Prevention and Control of DiseaseFrom EverandKey Questions in Clinical Farm Animal Medicine, Volume 3: Therapy, Prevention and Control of DiseaseTanmoy RanaNo ratings yet
- Wavy Changes in The Whiskers of Domestic Cats Are Correlated With Feline Leukemia Virus InfectionDocument7 pagesWavy Changes in The Whiskers of Domestic Cats Are Correlated With Feline Leukemia Virus InfectionCamila QuidelNo ratings yet
- Veterinary Internal Medicne - 2009 - Gleich - Hematology and Serum Biochemistry of Feline Immunodeficiency Virus InfectedDocument7 pagesVeterinary Internal Medicne - 2009 - Gleich - Hematology and Serum Biochemistry of Feline Immunodeficiency Virus InfectedCamila QuidelNo ratings yet
- Javma-Javma 228 3 371Document6 pagesJavma-Javma 228 3 371Camila QuidelNo ratings yet
- Veterinary Internal Medicne - 2017 - Levy - Performance of 4 Point of Care Screening Tests For Feline Leukemia Virus andDocument6 pagesVeterinary Internal Medicne - 2017 - Levy - Performance of 4 Point of Care Screening Tests For Feline Leukemia Virus andCamila QuidelNo ratings yet
- Dunham 2008Document23 pagesDunham 2008Camila QuidelNo ratings yet
- Estillero, Mari Antoinette - SPED 104 - Output Week 1 and 2Document4 pagesEstillero, Mari Antoinette - SPED 104 - Output Week 1 and 2Mari Antoinette EstilleroNo ratings yet
- OlivierDocument481 pagesOlivierVetriSelvan Nagarajan0% (1)
- A Case of Mitral StenosisDocument29 pagesA Case of Mitral StenosisShadab KamalNo ratings yet
- Drug Profile: C H FN ODocument2 pagesDrug Profile: C H FN Ohamed razaNo ratings yet
- Nutrition Concepts and Controversies 14Th Edition Sizer Test Bank Full Chapter PDFDocument53 pagesNutrition Concepts and Controversies 14Th Edition Sizer Test Bank Full Chapter PDFinlawbast.ileir100% (12)
- A Guide To Performing Quality Assured Diagnostic SpirometryDocument28 pagesA Guide To Performing Quality Assured Diagnostic SpirometryLaura MoiseNo ratings yet
- Genetic Factors Associated With Periodontal DiseasesDocument50 pagesGenetic Factors Associated With Periodontal DiseasesDrKrishna DasNo ratings yet
- Discovery and Development of Nucleoside and Nucleotide Reverse-Transcriptase InhibitorsDocument8 pagesDiscovery and Development of Nucleoside and Nucleotide Reverse-Transcriptase Inhibitorsolivia523No ratings yet
- 2016 ATC Index With DDDDocument317 pages2016 ATC Index With DDDSi CentilNo ratings yet
- TechTalk - August2010 Clump in EDTA Tubes PDFDocument2 pagesTechTalk - August2010 Clump in EDTA Tubes PDFARIF AHAMMED PNo ratings yet
- Brachycera 08Document27 pagesBrachycera 08Agustin IskandarNo ratings yet
- Lymphatic Drainage of Head and NeckDocument19 pagesLymphatic Drainage of Head and NeckKelly Yeow100% (1)
- Occupational DiseasesDocument10 pagesOccupational DiseasesAbhishek tiwariNo ratings yet
- E-Patients White PaperDocument126 pagesE-Patients White PaperPatients Know BestNo ratings yet
- Nursing Care Plan Sheet SampleDocument3 pagesNursing Care Plan Sheet Sampledbryant0101100% (5)
- Web 1 - Dr. Wahyuni Indawati, Sp. A (K) - Diagnosis and Management of Allergic Rhinitis in ChildrenDocument27 pagesWeb 1 - Dr. Wahyuni Indawati, Sp. A (K) - Diagnosis and Management of Allergic Rhinitis in Childrenbungkang elsaNo ratings yet
- Dog Parks in Hume City - Feasibility StudyDocument32 pagesDog Parks in Hume City - Feasibility Studykd.prawestiNo ratings yet
- Chemotherapy - Alkylating AgentsDocument33 pagesChemotherapy - Alkylating AgentsNolan100% (1)
- Homosexuality: A History (From Ancient Greece To Gay Liberation) Vern L. BulloughDocument209 pagesHomosexuality: A History (From Ancient Greece To Gay Liberation) Vern L. Bulloughhugoprais100% (1)
- Dea 2Document15 pagesDea 2Dea Hadfina Kartika SariNo ratings yet
- Management of Alveolar Ridge ResorptionDocument29 pagesManagement of Alveolar Ridge Resorptionفواز نميرNo ratings yet
- Principles of Medical Technology Practice 1 Midterm TranseesDocument19 pagesPrinciples of Medical Technology Practice 1 Midterm TranseesSean Rafael LedesmaNo ratings yet
- CVS Specialty Pharmacy Distribution Drug ListDocument19 pagesCVS Specialty Pharmacy Distribution Drug Listdp4mNo ratings yet
- Gadtc PHCDocument246 pagesGadtc PHCshawnNo ratings yet
- The Respiratory System I.: Subject: Subject TeacherDocument4 pagesThe Respiratory System I.: Subject: Subject TeacherHyacinth FarinasNo ratings yet
- Down's Syndrome Invetigsatory ProjectDocument23 pagesDown's Syndrome Invetigsatory ProjectKrithishNo ratings yet