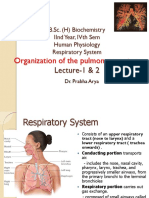
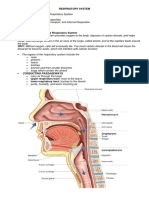
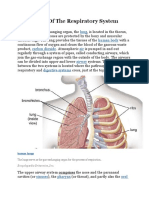
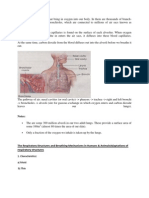
Download as pdf or txt
You might also like
- Cells AliveDocument13 pagesCells Aliveapi-3025396180% (1)
- GoldWingGL1800 (2020) CompressedDocument316 pagesGoldWingGL1800 (2020) Compressedchan swantananantNo ratings yet
- BC (H) - IV-Human Physiology, Respiratory System-1 & 2Document45 pagesBC (H) - IV-Human Physiology, Respiratory System-1 & 2sidra nazirNo ratings yet
- For Other Uses, See - "Lunged" Redirects Here. For Other Uses, SeeDocument12 pagesFor Other Uses, See - "Lunged" Redirects Here. For Other Uses, Seefifa92_piscesNo ratings yet
- Anatomy of Respiratory System in Poultry: ArticleDocument8 pagesAnatomy of Respiratory System in Poultry: ArticleJuwita Tabeta DamayantiNo ratings yet
- Breathing and Exchange of GasesDocument17 pagesBreathing and Exchange of GasesArjun ChristopherNo ratings yet
- ApbioprojectDocument5 pagesApbioprojectAshleyNo ratings yet
- Physiology of The Respiratory System in BirdsDocument3 pagesPhysiology of The Respiratory System in BirdsScribdTranslationsNo ratings yet
- Respiratory System of Bird FinalDocument10 pagesRespiratory System of Bird FinalMonjurul Haque MilonNo ratings yet
- Accessory Respiratory Organ - FishDocument8 pagesAccessory Respiratory Organ - Fishkaushik mallickNo ratings yet
- Respiratory Diseases of Parrots - Anatomy, Physiology, Diagnosis and Treatment - Crosta, L. (2021)Document22 pagesRespiratory Diseases of Parrots - Anatomy, Physiology, Diagnosis and Treatment - Crosta, L. (2021)Veronica Hdez FloresNo ratings yet
- Theory of LungDocument7 pagesTheory of LungAndres Dario R LozNo ratings yet
- Anatomic and Physiologic OverviewDocument8 pagesAnatomic and Physiologic OverviewJoseph King MacaranasNo ratings yet
- Respiratory AssessmentDocument12 pagesRespiratory AssessmentAmanda ScarletNo ratings yet
- Breathing and Exchange of GasesDocument8 pagesBreathing and Exchange of GasesgoswamiuwuNo ratings yet
- Respiratory SystemDocument6 pagesRespiratory SystemQK CLARONo ratings yet
- Respiratory SystemDocument5 pagesRespiratory SystemJay QuilnetNo ratings yet
- Inhaled Anesthesia For BirdsDocument18 pagesInhaled Anesthesia For BirdsBianca PaludetoNo ratings yet
- Respiratory System: Anatomy & PhysiologyDocument111 pagesRespiratory System: Anatomy & PhysiologyJape GarridoNo ratings yet
- The Design of The Respiratory System: Lung Human Body Carbon Dioxide Air Airway Digestive Systems LarynxDocument5 pagesThe Design of The Respiratory System: Lung Human Body Carbon Dioxide Air Airway Digestive Systems LarynxAndrewNo ratings yet
- Human Anatomy Reading ComprehensionDocument16 pagesHuman Anatomy Reading ComprehensionRhams BairullaNo ratings yet
- Respiratory PhysiologyDocument15 pagesRespiratory PhysiologySahil SalluNo ratings yet
- Respiratory SystemDocument53 pagesRespiratory SystemRotan CirebonNo ratings yet
- Respiratory System PreDocument11 pagesRespiratory System PrecontehamosaNo ratings yet
- Pathophysiology of Respiratory System Feghiu I., Tacu L. Lutan VDocument55 pagesPathophysiology of Respiratory System Feghiu I., Tacu L. Lutan VLunguVictoriaNo ratings yet
- PAin and Oxygenation HandoutsPRELIMSDocument7 pagesPAin and Oxygenation HandoutsPRELIMSJhoanna de ChavezNo ratings yet
- PDBreathing and Exchange of Gases Class 11 Notes CBSE Biology Chapter 17FDocument15 pagesPDBreathing and Exchange of Gases Class 11 Notes CBSE Biology Chapter 17Fgandhitirth147No ratings yet
- The Design of The Respiratory SystemDocument21 pagesThe Design of The Respiratory Systemvonneivan26No ratings yet
- Our Lungs Are The Organs That Bring in Oxygen Into Our BodyDocument17 pagesOur Lungs Are The Organs That Bring in Oxygen Into Our BodySaidatul Atyah Mohd ApendaiNo ratings yet
- Supplementary Material 3.1 Basic Concepts and Assessment (N) - 2Document13 pagesSupplementary Material 3.1 Basic Concepts and Assessment (N) - 2Kent Borro CantomayorNo ratings yet
- Kebo114 PDFDocument12 pagesKebo114 PDFLukesh SadhNo ratings yet
- Gas Exchange Written ReportDocument11 pagesGas Exchange Written ReportEllanny Gloria100% (1)
- Bilology XII 2014 CH - 14 RespirationDocument16 pagesBilology XII 2014 CH - 14 RespirationMuhammad UsmanNo ratings yet
- Problems With OxygenationDocument92 pagesProblems With OxygenationEbiNo ratings yet
- Anatomy and Physiology Power PointDocument8 pagesAnatomy and Physiology Power PointJoanna Diongson GonzalesNo ratings yet
- Respiratory SystemDocument7 pagesRespiratory SystemMark Andrew CabaleNo ratings yet
- Noor RespiratoryDocument45 pagesNoor RespiratoryHond HayNo ratings yet
- Bronchial AsthmaDocument21 pagesBronchial AsthmaMoonNo ratings yet
- OxygenationDocument20 pagesOxygenationKhie-An OcampoNo ratings yet
- The Respiratory System or Pulmonary SystemDocument53 pagesThe Respiratory System or Pulmonary Systemfharidaismail47No ratings yet
- Human Respiratory System - Britannica Online EncyclopediaDocument31 pagesHuman Respiratory System - Britannica Online EncyclopediaAlina AluculeseiNo ratings yet
- Respiratory and Circulatory SystemDocument10 pagesRespiratory and Circulatory Systemainabukhari20No ratings yet
- 16 Respiratory SystemDocument40 pages16 Respiratory SystemchromaNo ratings yet
- Respiratory System s2 2019Document56 pagesRespiratory System s2 2019noviantyramadhani12No ratings yet
- Respiratory SystemDocument42 pagesRespiratory SystemSohailriazNo ratings yet
- Respiratory System: A) The Conducting Portion, Which Consists of The Nasal CavitiesDocument7 pagesRespiratory System: A) The Conducting Portion, Which Consists of The Nasal CavitiesSalmah Alimah FitriNo ratings yet
- Resumen Primera Parte. FisiologíaDocument4 pagesResumen Primera Parte. FisiologíaFatima AzaleansNo ratings yet
- Respiratory System A. BDocument5 pagesRespiratory System A. BPhoebe Kates Dela CruzNo ratings yet
- Breathing and Exchange Notes-2Document11 pagesBreathing and Exchange Notes-2manshibhaskar09No ratings yet
- Handout Respiratory System 2022 2023Document3 pagesHandout Respiratory System 2022 2023reece sayamNo ratings yet
- AICE Bio Notes Pages 186-190: Gas ExchangeDocument2 pagesAICE Bio Notes Pages 186-190: Gas ExchangeSanzida HossainNo ratings yet
- Materi BingDocument17 pagesMateri BingCindy Ari SantiNo ratings yet
- What Protections Does The Upper Airway Have For The Lower Airway?Document3 pagesWhat Protections Does The Upper Airway Have For The Lower Airway?Paul JacksonNo ratings yet
- Indah Yulianti - 012011041 - Kep A1'20 - Resume Respiratory SystemDocument6 pagesIndah Yulianti - 012011041 - Kep A1'20 - Resume Respiratory SystemIndah YuliantiNo ratings yet
- The Lung Spech NewDocument11 pagesThe Lung Spech NewyunitaNo ratings yet
- Anatomy and Physiology of Respiratory SystemDocument34 pagesAnatomy and Physiology of Respiratory SystemSamson SeiduNo ratings yet
- Chapter 13 The Respiratory SystemDocument7 pagesChapter 13 The Respiratory Systemqueen nurseNo ratings yet
- Iv. Anatomy and Physiology of The Human Respiratory SystemDocument5 pagesIv. Anatomy and Physiology of The Human Respiratory SystemJenny Vi CodenieraNo ratings yet
- Anatomy of The RespiratoryDocument8 pagesAnatomy of The Respiratoryjoanna gurtizaNo ratings yet
- Thoracic and Coracoid Arteries In Two Families of Birds, Columbidae and HirundinidaeFrom EverandThoracic and Coracoid Arteries In Two Families of Birds, Columbidae and HirundinidaeNo ratings yet
- HTGD 90117 - Aa - Instruction For Lube OilDocument22 pagesHTGD 90117 - Aa - Instruction For Lube Oilabdikariev.aybekNo ratings yet
- MP 12th Five Year PlanDocument599 pagesMP 12th Five Year Planphilodog100% (2)
- Urban Transit Scheduling: Framework, Review and Examples: AbstractDocument20 pagesUrban Transit Scheduling: Framework, Review and Examples: AbstractRichie Jeet SinhaNo ratings yet
- Devil and Angel in The Renin-Angiotensin System ACE-angiotensin II-AT1 Receptor Axis vs. ACE2-angiotensin - (1-7) - Mas Receptor AxisDocument4 pagesDevil and Angel in The Renin-Angiotensin System ACE-angiotensin II-AT1 Receptor Axis vs. ACE2-angiotensin - (1-7) - Mas Receptor AxisLevente BalázsNo ratings yet
- Classroom of The Elite Volume ODocument271 pagesClassroom of The Elite Volume OPrashant ShuklaNo ratings yet
- Geared Motor Device 100/130V E1/6-T8Document2 pagesGeared Motor Device 100/130V E1/6-T8seetharaman K SNo ratings yet
- Chapter 2.1Document152 pagesChapter 2.1Naoto MikaNo ratings yet
- Junos Release Notes 12.3r11 PDFDocument390 pagesJunos Release Notes 12.3r11 PDFHellman Roman Pinzon GarciaNo ratings yet
- Chim Chim WorkoutDocument5 pagesChim Chim Workoutretro100% (5)
- Course Outline Jan 2012Document4 pagesCourse Outline Jan 2012Urim ParkNo ratings yet
- T.l.e9 T1 Tools and Equipment Needed in Preparing AppetizersDocument38 pagesT.l.e9 T1 Tools and Equipment Needed in Preparing AppetizersRedginald CalderonNo ratings yet
- Isr4321 k9 DatasheetDocument6 pagesIsr4321 k9 DatasheetSail DMNo ratings yet
- FILE 20210720 230205 Ôn-Tập-TACNDocument9 pagesFILE 20210720 230205 Ôn-Tập-TACNVăn KhánhNo ratings yet
- Math 5 Q3 Week 7Document11 pagesMath 5 Q3 Week 7Cristina Morante100% (2)
- BF 01202562Document3 pagesBF 01202562Josiel Nasc'mentoNo ratings yet
- Marine Propulsion System Failures-A Review: Marine Science and EngineeringDocument14 pagesMarine Propulsion System Failures-A Review: Marine Science and EngineeringDavid NuñezNo ratings yet
- Definium Amx 700Document240 pagesDefinium Amx 700محمدعبدالخالق العلوانيNo ratings yet
- Black KnightDocument3 pagesBlack KnightMattNo ratings yet
- Pamantasan NG Lungsod NG Maynila Intramuros, ManilaDocument2 pagesPamantasan NG Lungsod NG Maynila Intramuros, ManilaWacko AsahanNo ratings yet
- Hazards and Risks in The KitchenDocument30 pagesHazards and Risks in The KitchenCynthia Lopez100% (1)
- Lesson #2: To Buy or Not Buy?: Bodily Self-PreservationDocument2 pagesLesson #2: To Buy or Not Buy?: Bodily Self-PreservationGilyn NaputoNo ratings yet
- Water Needs and Hydration For Cats and Dogs - Brain M.zanghiDocument9 pagesWater Needs and Hydration For Cats and Dogs - Brain M.zanghiann nnNo ratings yet
- Ccna Class NotesDocument262 pagesCcna Class Notessyed latheefNo ratings yet
- 4Document2 pages4waar lockNo ratings yet
- Haryana With CommentsDocument148 pagesHaryana With Commentsmanojkumartomar7513No ratings yet
- List of AGMs Promoted As GMs W.E.F. 25.06.2023Document2 pagesList of AGMs Promoted As GMs W.E.F. 25.06.2023Abhishek ChoubeyNo ratings yet
- 2101 ManDocument110 pages2101 ManValentin soliñoNo ratings yet
- FM 8900 SDocument136 pagesFM 8900 SRomeish KelaartNo ratings yet